| |
Karrerulina conversa (Grzybowski, 1901)
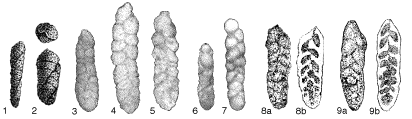
Fig. 116. 1-2. Type figures of Gaudryina conversa from Grzybowski (1901);
3-5. Type figures of Karrerulina bortonica from Finlay (1940);
5-7. Type figures of Karrerulina aegra from Finlay (1940);
8. Holotype of Karreriella indigena from Mjatliuk (1970);
9. Holotype of Karreriella pokornyi from Mjatliuk (1970).
ORIGINAL DESIGNATION: Gaudryina conversa Grzybowski, 1901
TYPE REFERENCE: Grzybowski, J., 1901, Otwornice warstw inoceramowych okolicy Gorlic. Rozprawy Wydziału Matematyczno-Przyrodniczego, Akademia Umiejętności w Krakowie, serya 2, 41, 219-286. pl. 8, figs. 15, 16. See also: Kaminski, M.A. & Geroch, S., 1993. A revision of foraminiferal species in the Grzybowski Collection. Grzybowski Foundation Special Publication no. 1, 239-323 [Lectotype].
TYPE SPECIMEN: Not originally designated. Grzybowski listed three samples containing specimens of Gaudryina conversa: Two samples from well Kalinka-1, (120 m and 244 m depth) drilled in the village of Ropica Polska, Poland, and one sample from well Bartne-1 (315 m) drilled in the village of Bartne. The two samples from the Kalinka well are preserved in the Grzybowski Collection, housed in the Jagiellonian University, Kraków, but our examination of these samples yielded no specimens of this species. The lectotype specimen designated by Kaminski & Geroch (1993; pl. 13, figs. 5a,b) was found in the Grzybowski Collection in a vial labelled "Gaudryina sp. beta II14". It displays a typical, well-developed biserial part, and necklike extension of the terminal aperture. The specimen is deposited in the Geological Museum of the Jagiellonian University, Kraków, and is registered as UJ-133-P, 2/105b.
TYPE LEVEL: Eocene. Grzybowski's Sample Kalinka-1, 120 m is from the upper Eocene A. latus Zone of Geroch & Nowak (1984). Sample Kalinka-1, 244 m is from the middle Eocene A. amplectens Zone of Geroch & Nowak (1984). Grzybowski's sample from Bartne (which corresponds to our neotype locality) was probable lower Eocene based on the absence of Paleocene and middle Eocene index species in his faunal list (Grzybowski, 1901, p. 243).
TYPE LOCALITY: Neotype locality (designated herein): red clays in a stream bed in the village of Bodaki, Poland, Magura Unit of the Polish Carpathians. This is one of Grzybowski's original localities (Locality #108 of Grzybowski, 1901) that is still accessible for collecting.
DIAGNOSTIC FEATURES: Test free, with an initial trochospiral portion of 3-4 whorls with 4 or 5 chambers per whorl, followed by one or two whorls with triserial arrangement and finally by a biserial stage of varying length which may be twisted along the long axis of the test. Mature specimens consist of 9-10 whorls and the test may be arched to one side. The initial trochospiral portion makes up about one-third the length of the test and is generally round in cross-section. The biserial portion is generally flattened and may possess a straight or lobate periphery. Sutures in the multiserial part are indistinct, but are usually incised in the biserial part. Chambers increase in size rapidly in the multiserial part, slowly in the biserial part. The ultimate chamber may be slightly more inflated and sometimes assumes a staggered orientation atop the penultimate chamber. Chamber interiors simple, sac-like, joined by tubular passages. Aperture terminal, rounded, at the end of a short neck. Wall medium to finely agglutinated, made of quartz grains with much cement.
SIZE: Dimensions of Lectotype: length 0.68 mm.
SUSPECTED SYNONYMS:
Gaudryina apicularis Cushman. Cushman, J.A., 1911, U.S. Nat. Mus. Bull. 71, p. 68, figs 110a, 110b. [Recent].
Karreriella (Karrerulina) aegra Finlay. Finlay, H.J., 1940, Trans. Proc. Royal Soc. New Zealand, 69, p. 451, pl. 62, figs. 21-22. ["Danian and lower Bortonian", New Zealand].
Karreriella (Karrerulina) bortonica Finlay. Finlay, H.J., 1940, ibid. p. 451, pl. 62, figs. 18-20 ["Bortonian", New Zealand].
Karreriella indigena Mjatliuk. Mjatliuk, E.V., 1970, Trudy VNIGRI 282, p. 116-117, pl. 34, figs. 10a-14b [upper Eocene, Ukrainian Carpathians].
Karreriella pokornyi Mjatliuk. Mjatliuk, E.V., 1970, Trudy VNIGRI 282, p. 117-118, pl. 34, figs. 15a-17 [upper Eocene, Ukrainian Carpathians].
OBSERVED OCCURRENCES: In the Alpine regions, this species has been recorded as Plectina conversa from the Eocene of the western Polish Carpathians (Geroch, 1960), as Karreriella conversa and Karreriella apicularis from the Eocene of the Carpathian units in eastern Poland (Jurkiewicz, 1967), as Plectina conversa from the Senonian to Eocene of the Polish Carpathians by Morgiel & Szymakowska (1978), and as Karrerulina conversa from the Paleocene-Eocene of the Dukla Unit by Bąk (2004), who also noted a Karrerulina spp.-dominated biofacies in the lower Eocene. Mjatliuk (1970) reported this form as Karreriella indigena and Karreriella pokornyi from the upper Eocene of the Lower Bystritsa Formation in the Ukrainian Carpathians. Verdenius & Van Hinte (1983) reported K. conversa from the Eocene of Norwegian-Greenland Sea DSDP sites.
Cushman & Renz (1946) reported this species as Gaudryina bentonensis (Carmen) from the Lizard Springs Formation of Trinidad. Specimens from the Eocene Navet Formation, San Fernando, Trinidad are housed in a slide labelled "Gaudryina cf. bentonensis" in the Cushman Collection.
The oldest reported verified occurrence of K. conversa is from the Upper Cretaceous of DSDP Site 367, off West Africa (Krasheninnikov & Pflaumann, 1977). At Site 367, this species occurs in the lateral equivalent of the Plantagenet Formation in an assemblage containing Uvigerinammina jankoi.
We have observed Karrerulina conversa in the Paleocene to Eocene of the Central North Sea and Labrador Margin [originally reported by Gradstein & Berggren (1981) as Karreriella apicularis] the Eocene of ODP Hole 647A in the southern Labrador Sea, the Eocene of ODP Hole 643 in the Norwegian Sea, the Paleocene-lower Eocene Lizard Springs Formation, in the middle Eocene Navet Formation, and in the Oligocene Cipero Formation in Trinidad, in the Paleocene to Eocene of Morocco, the Paleocene of Zumaya, Spain, in the Paleocene Kamalapuram Formation of the Cauvery Basin, India, in the Oligocene at ODP Site 1148 in the South China Sea (Kuhnt et al. 2002), and in the upper Miocene at ODP Site 646 off the southern tip of Greenland.. We have also observed K. conversa in the lower Eocene (S. patagonica Zone) of Burdwood Bank in the South Atlantic in dredge samples recovered by the RSS Discovery II and the RSS Scoresby.
In the central North Sea, K. conversa disappears on average in the middle to upper Eocene R. amplectens Zone above the North Sea Log Marker E. It was not observed in younger strata. On the Northern Grand Banks and Labrador Margin, K. conversa disappears on average in the middle Eocene P. aff. paucicostata Zone, but may range into the overlying R. amplectens Zone (middle to upper Eocene). The oldest occurrence in offshore eastern Canada is in the upper Maastrichtian A. mayaroensis Zone of the Indian Harbour M-52 well. In Zumaya, Ortiz (1995) found a K. conversa acme immediately above the Paleocene/Eocene boundary, coincident with a peak in Glomospira charoides. In the North Sea, Poland, and in Morocco, it forms an acme in the upper part of the lower Eocene, while at ODP Site 647 a maximum in K. conversa is observed in the uppermost part of the lower/middle Eocene Glomospira facies. Moderrn representatives of the genus Karrerulina are common on the abyssal plains, in environments characterised by very low organic carbon flux. Its acmes in Eocene strata are therefore linked to changes in the trophic continuum and reflect more oligotrophic conditions, typically characterised by the deposition of red shales in the Atlantic and Western Tethys.
KNOWN STRATIGRAPHIC RANGE: Predominantly Paleocene to Oligocene, with rare occurrences in the Miocene to Recent, and in the Senonian (Upper Cretaceous). Reports of the species from the Turonian (Hanzlíková, 1953, 1972; Malata & Oszczypko, 1990) and Albian (Pflaumann, 1964) require verification.
BATHYMETRY: upper bathyal to abyssal.
REMARKS: Karrerulina conversa differs from Karrerulina horrida and Karrerulina coniformis in possessing a well-developed twisted biserial part, that in mature specimens comprises over half the length of the test. Individual immature specimens may be difficult to distinguish from K. horrida, however, in large populations specimens with biserial parts always occur.
Grzybowski's description of Gaudryina conversa is brief and inaccurrate regarding the position of the aperture. This was first noted by Pokorny (1953) who placed this form in "Plectina conversa Jedlitschka, 1935 (non Grzybowski)" because of its terminal aperture on a distinct neck. Mjatliuk regarded Grzybowski's drawing of Gaudryina conversa to be "incomprehensible", and chose to abandon the specific designation. Instead, she described the new species Karreriella indigena, under which she synonymized Plectina conversa of Geroch (1960). Although the cotypes of Gaudryina conversa are missing from the Grzybowski Collection, we believe it is unnecessary to abandon this name or regard the species as nomen dubium. Kaminski & Geroch (1993) selected a lectotype specimen that was found in the Grzybowski Collection in a vial labelled "Gaudryina sp. beta II14". This specimen displays a typical, well-developed biserial part, and is possibly the plesiotype of Grzybowski (1901; pl. 7, fig. 15), but this specimen is broken at the anterior end. Our lectotype (pl. 116, fig. 7) consists of an initial trochospire of about four whorls with four chambers to a whorl, one whorl of three chambers, and four twisted pairs of biserial chambers.
Because samples from Grzybowski's wells are no longer available, we have designated a neotype locality from one of Grzybowski's original outcrop sections. The specimens from the Magura Unit of the Polish Carpathians are strongly silicified, which allows the interior of the test to be viewed clearly using immersion oil. The specimens from the neotype locality consist mainly of smaller specimens, some with only one or two pairs of biserial chambers, others ending in a triserial chamber arrangement. Mjatliuk (1970) also reported frequent immature specimens in her material from the eastern Carpathians, and this is also seen in assemblages from the North Sea. Specimens from the North Sea vary mainly in the coarseness of agglutinated grains and the nature of the test outline, with slender specimens less common than wider specimens with lobate outlines.
Karrerulina bortonica Finlay and Karrerulina aegra (regarded as a junior synonym of the former by Hornibrook et al. 1989) were described from the "Danian, Bortonian, and Kaiatian" of New Zealand. Hornibrook et al. 1989 reported the stratigraphical range of this form as upper Paleocene to mid-late Eocene. Karrerulina aegra was originally reported by Finley to be "somewhat smaller and more slender" than K. bortonica. We agree with Hornibrook in regarding both of Finlay's species to be synonymous, but we regard them as junior synonyms of K. conversa.
The considerable variation in the outline of the tests of the modern species Gaudryina apicularis Cushman from the deep North Atlantic, and comparison of Recent material to the original collection of H.B. Brady and the neotypes of K. conversa have led us to the conclusion that G. apicularis is a junior synonym. This synonymy explains the fact that it was often difficult to assign specimens from the Eocene of the North Sea to either K. conversa or "K. apicularis". Karreriella apicularis is supposedly less slender and straight-edged with a less twisted biserial part than K. conversa, but this distinction is not tenable in the material at our disposal. Recent specimens from the Nova Scotian continental rise possess well-developed, twisted biserial parts and inflated chambers. Specimens from the Hatteras Abyssal Plain are more slender and more finely agglutinated. The range in variation in size and coarseness of the wall observed in fossil specimens of K. conversa has a good analog in recent populations. The junior synonymy of K. apicularis considerably extends the known stratigraphic range of K. conversa, and alleviates the problem of deciding which specific name to use for Miocene specimens.
There are two Cretaceous taxa that may be confused with K. conversa, i.e. Gaudryina filiformis Berthelin, and Gerochammina lenis (Grzybowski). The former posseses an interiomarginal, slit aperture at the base of the last chamber, and often has a biserially twisted part of the test that is much longer than its rest Gaudryina filiformis occurs in mid-Cretaceous strata. It is not known if the wall cement is actually calcareous, as is typical for Gaudryina. The latter taxon, Gerochammina lenis, more closely resembles K. conversa, but has a less pronounced apertural neck, and a less well-developed biserial part (Hus, 1966, pl. 8, figs. 1-5; Neagu 1990, pl. 2, figs. 22-30.), and is typical for Upper Cretaceous, Senonian strata. We consider G. lenis to be ancestral to K. conversa, with the evolutionary status of Gaudryina filiformis unclear.
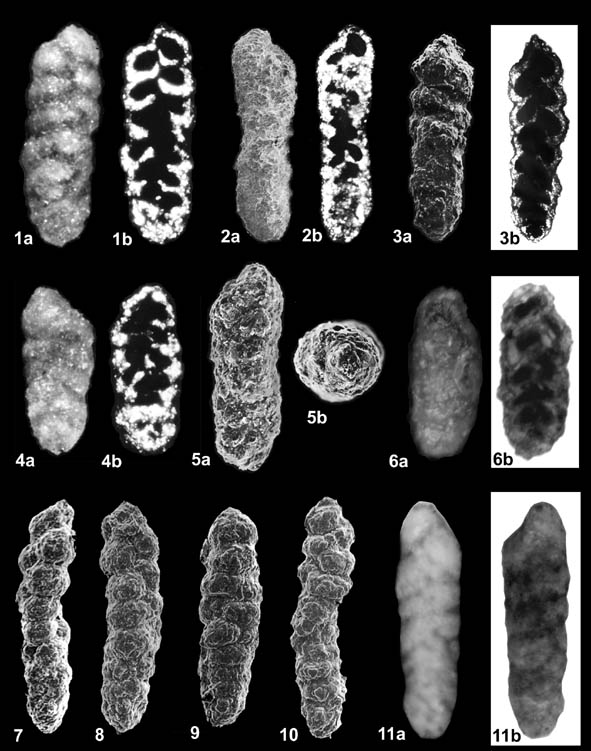
ILLUSTRATIONS: Plate 116 - Karrerulina conversa (Grzybowski)
Fig. 1a,b. Lectotype, Magura Unit of the Polish Carpathians, Ropica Polska, (UJ-133-P 2/105b, as "Gaudryina sp. beta II14") reflected and cross-polarised light; Fig. 2a,b. Krościenko Niżne, middle Eocene variegated clays, outcrop on the bank of the Wisłok River; Fig. 3a,b. Eocene, Magura Unit of the Polish Carpathians, variegated clays from a streambed in Bartne Poland (locality nr. 72 of Grzybowski (1901); Figs. 4a-5b. Paralectotypes, Magura Unit of the Polish Carpathians, Ropica Polska. 5b. apertural view; Fig. 6a,b. Upper Eocene, Nizhnebystritskaya svita, Ratunzova River, Ukraine, syntype of "Karreriella indigena Mjatliuk", reflected and transmitted light in immersion; Figs. 7-8. Middle Eocene, Labrador Margin, South Labrador N-79, 1980-1990m; Figs. 9-10. Lower Eocene, North Sea, Esso Norway 16/1-1 well, 7060'; Fig. 11a,b. Upper Eocene, Nizhnebystritskaya svita, S. Busovisko, Ukraine, syntype of "Karreriella pokornyi Mjatliuk", reflected light & in immersion.
|
|