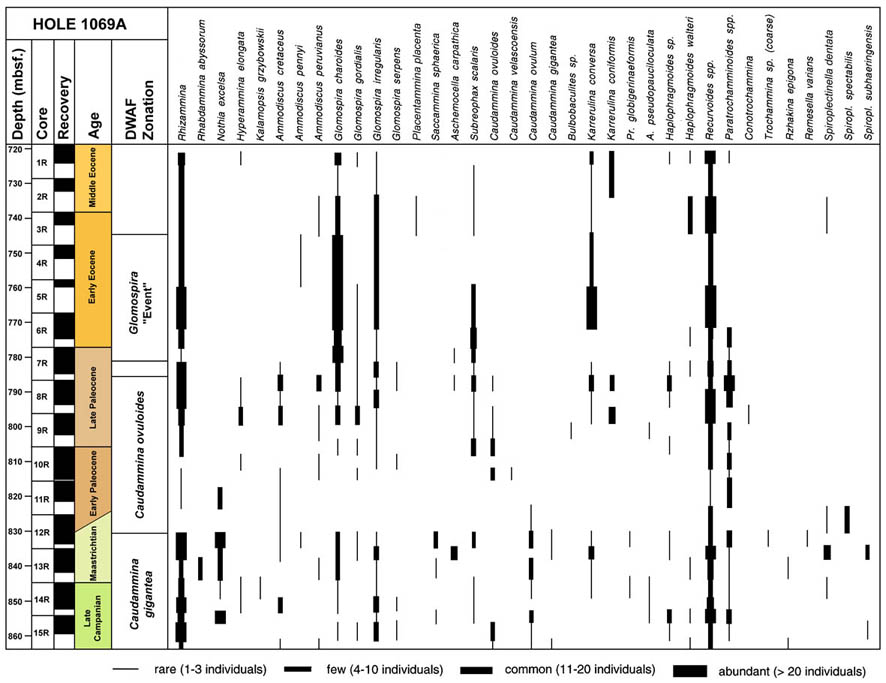
SUMMARY
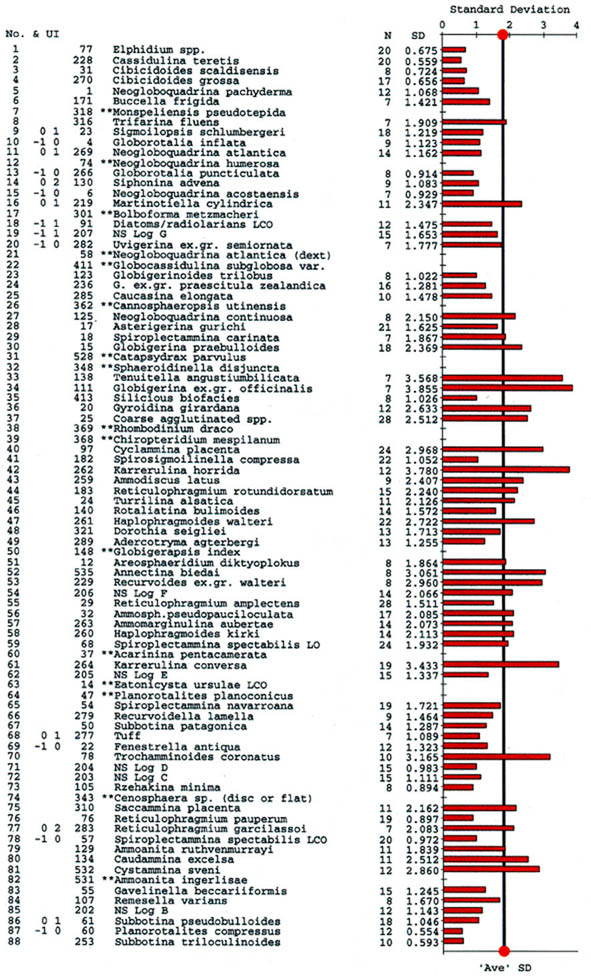
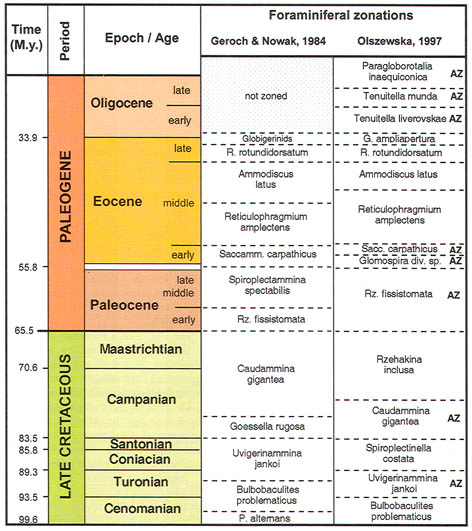
Zonal Schemes
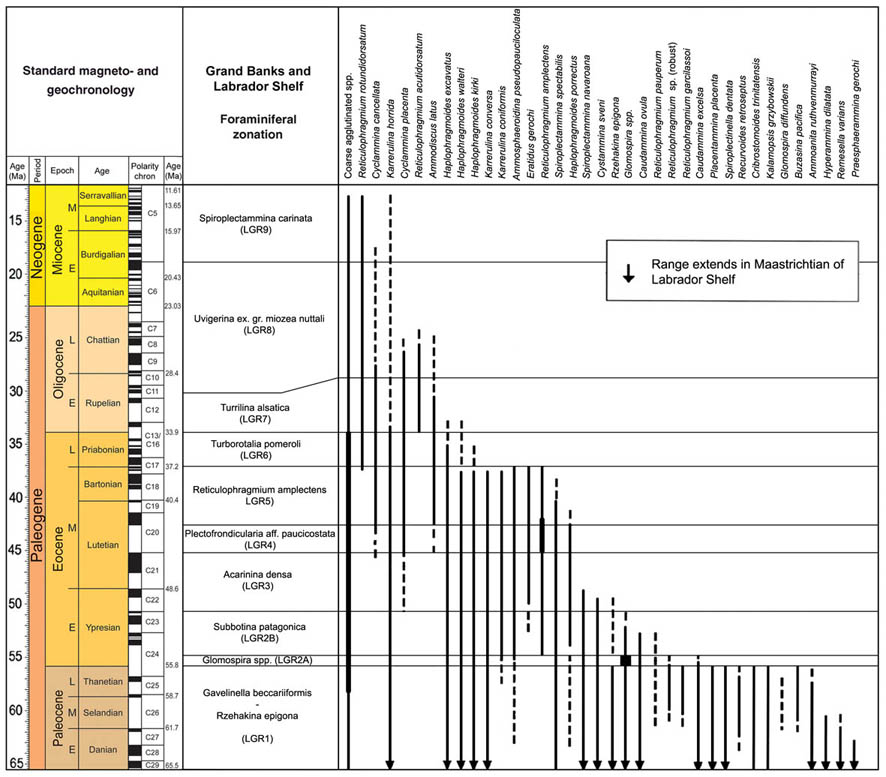
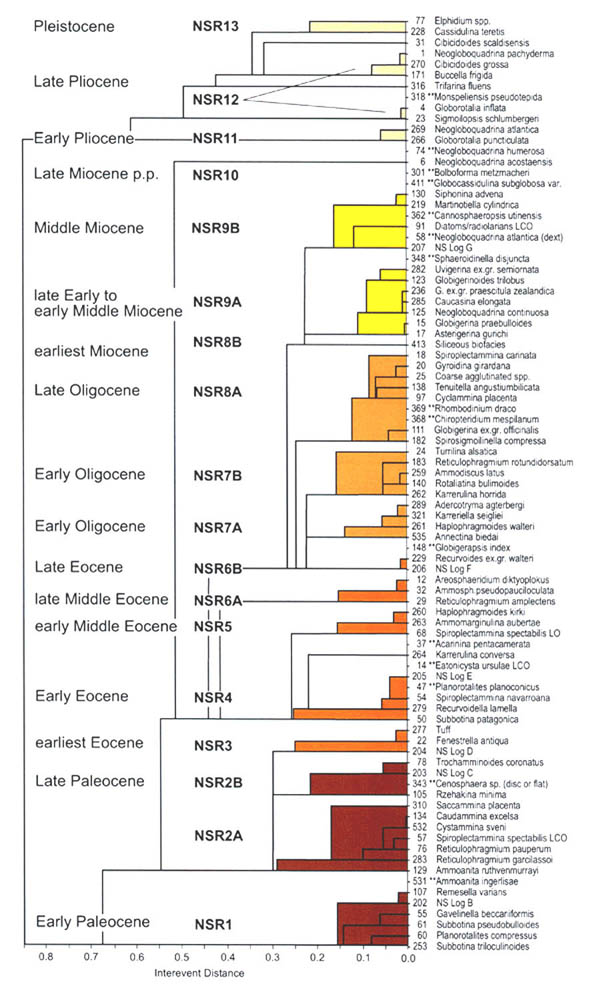
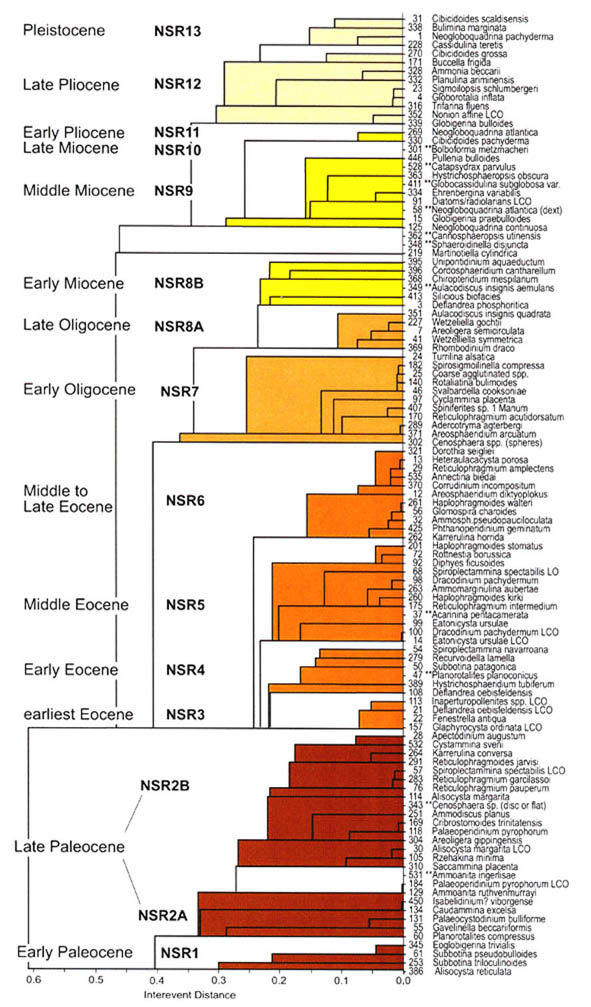
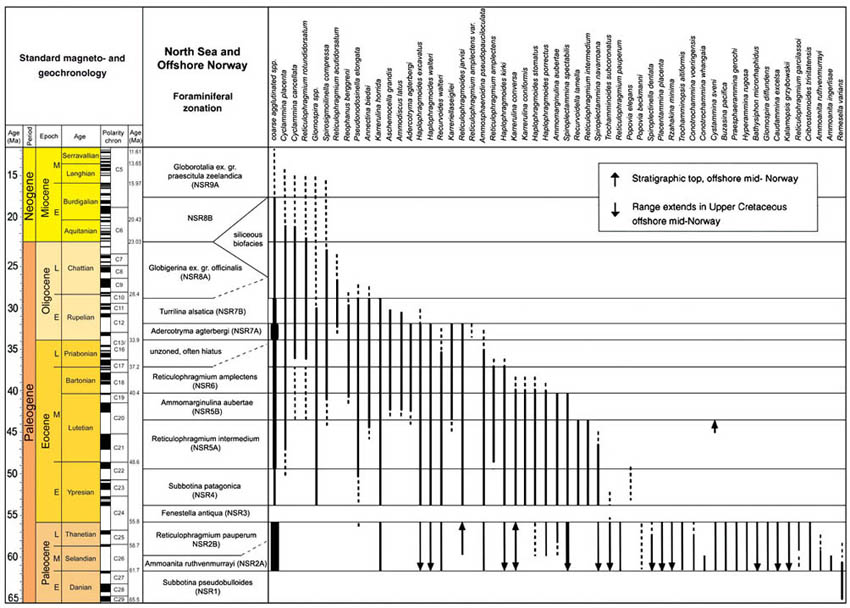
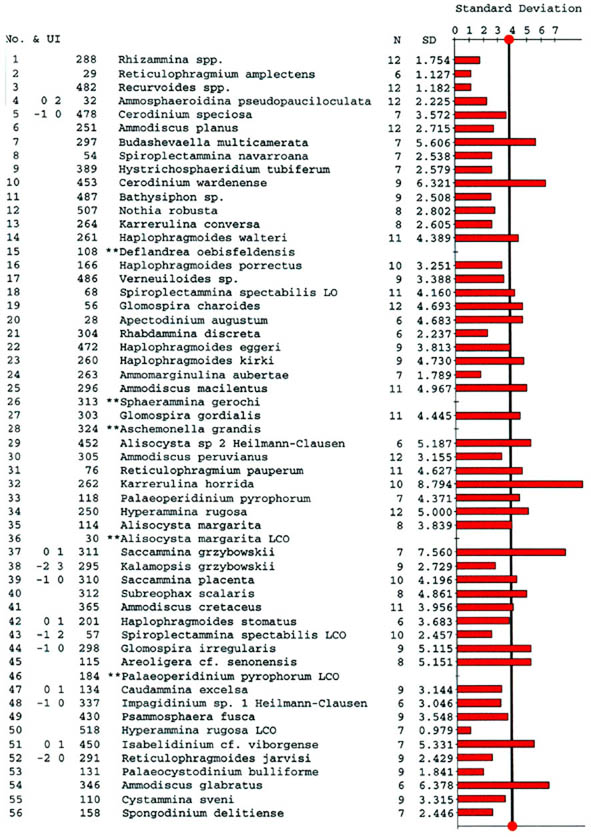
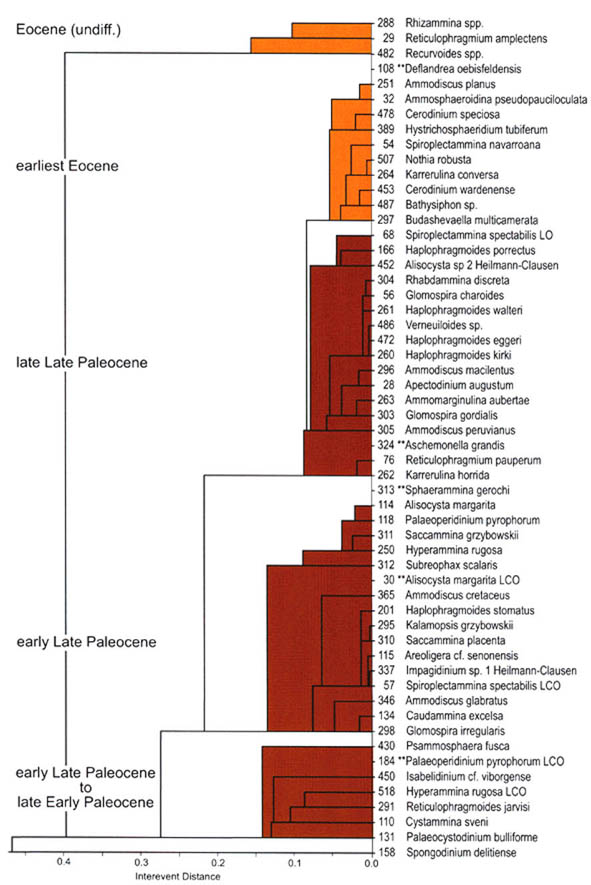
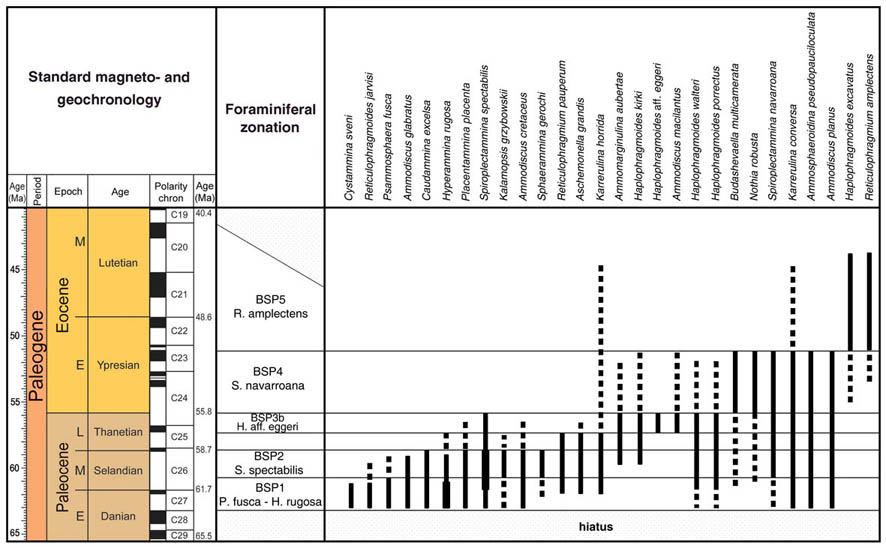
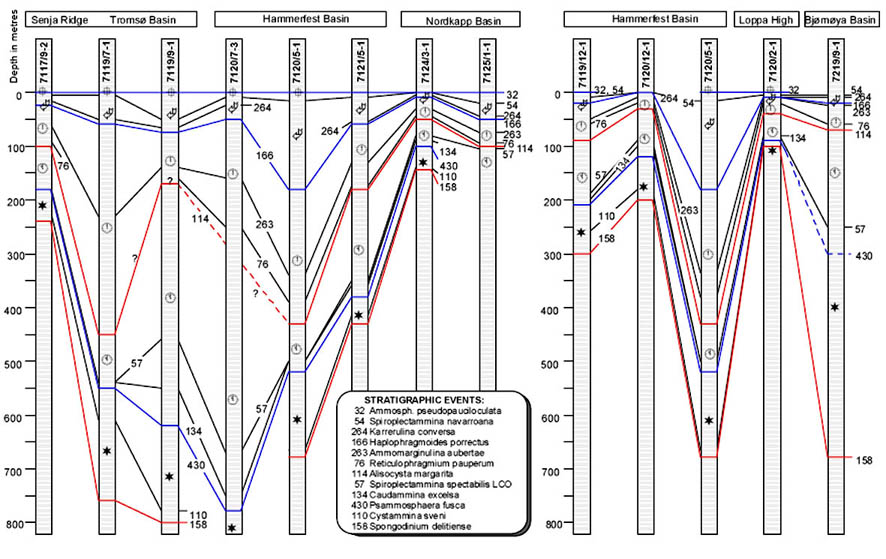
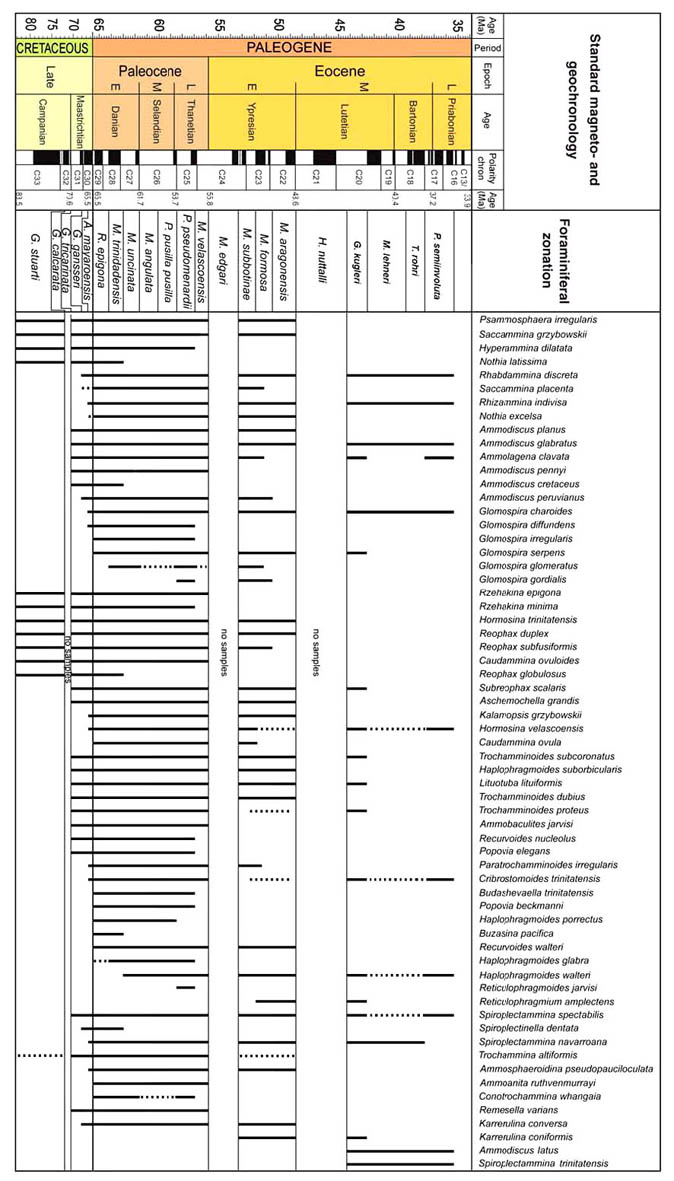
The preceeding discussion of the studied localities should provide an overview of the use of DWAF for constructing local stratigraphic zonations and for correlation of deep marine sequences. The main point that can be made is the fact that DWAF may be very useful for intra-basinal stratigraphical correlations. However, for the purpose of inter-regional correlation, environmental factors such as palaeobathymetry and the trophic continuum must be taken into account before we attempt long-distance correlations. The silled basins such as the Norwegian Sea, and to some extent the various flysch basins of the Alpine-Carpathian area have their own depositional histories and degrees of faunal endemism. Both the Geroch & Nowak "Tethyan" zonation and the Gradstein et al. "Boreal" zonations reflect basinal history, and have some events that are unique to a given region. However, there are also some interesting communalities between the Tethys and the Boreal basins that may be related to inter-basinal marine conections or to oceanic events of a more global nature. Examples of the latter are the Cretaceous/Paleocene boundary event, the Paleocene/Eocene boundary, and the Eocene/Oligocene boundary events. Each of these events resulted in important faunal changes among the DWAF in terms of extinctions and/or the occurrence of a particular biofacies. A summary of Paleogene zonal schemes from the western Tethys to the Arctic Ocean is given in figure 44.
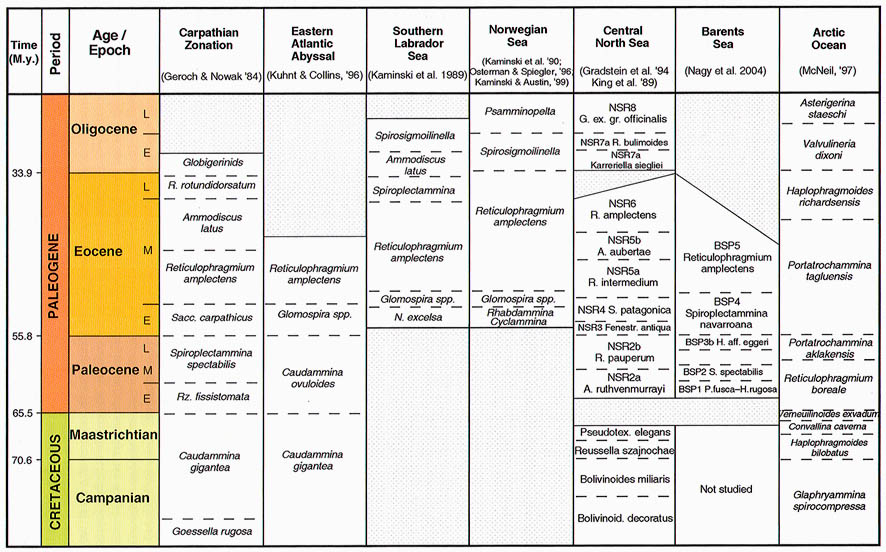
Figure 44. Summary of Paleogene zonal schemes described in different regions, arranged approximately from south to north.
Paleogene DWAF Acme Events in the Western Tethys and North Atlantic
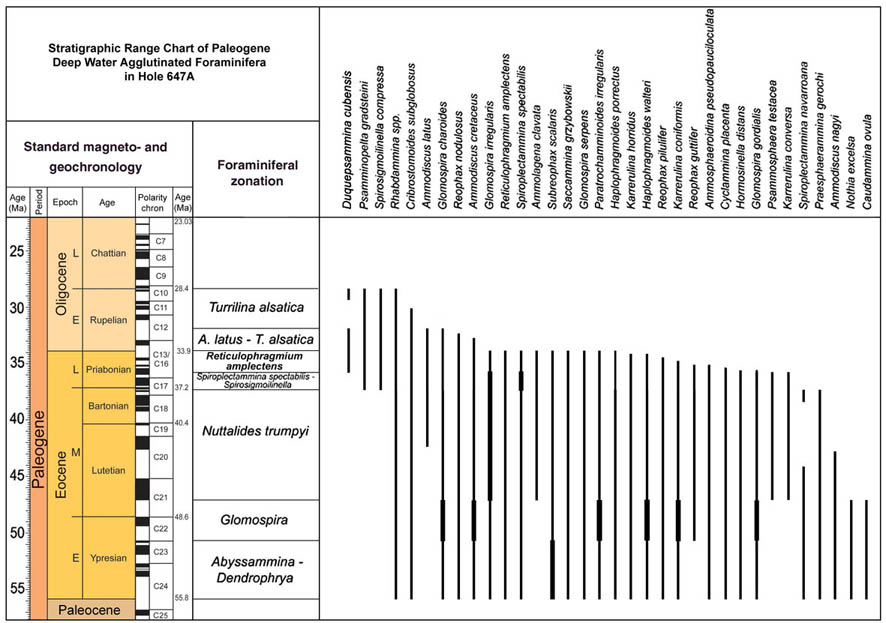
The traditional zonal schemes proposed for Tethyan and North Atlantic DWAF were based on first and last occurrences of characteristic taxa (e.g., Geroch & Nowak, 1984, King, 1989). Other zonal schemes have made use of a combination of first and last occurrences as well as abundance maxima (e.g., Olszewska, 1997). It is becoming increasingly apparent that the important oceanic events of the Paleogene have been an important instrument of benthic faunal change in the deep ocean. Quantitative analysis of Deep-water Agglutinated Foraminifera (DWAF) assemblages from key ODP sites in the North Atlantic reveal the presence of stratigraphically-significant abundance maxima, that may be useful for correlating sedimentary sequences deposited beneath the CCD. For the purpose of this summary, the DWAF record from ODP Hole 647A in the Labrador Sea was re-studied and abundances were recalculated by excluding calcareous benthic foraminifera (Fig. 45). This hole is a key locality, as it provides direct calibration of the DWAF biostratigraphy to the standard chronostratigraphy. A second important locality for the calibration of the acme events is the Contessa Road section, where several acme events are developed to a greater or lesser extent. These acmes, in turn can be traced to the eastern Atlantic ODP sites (Kuhnt & Collins, 1996; Kuhnt & Urquhart, 2001) and to the Polish Carpathians (Olszewska, 1997, Bąk, 2004).
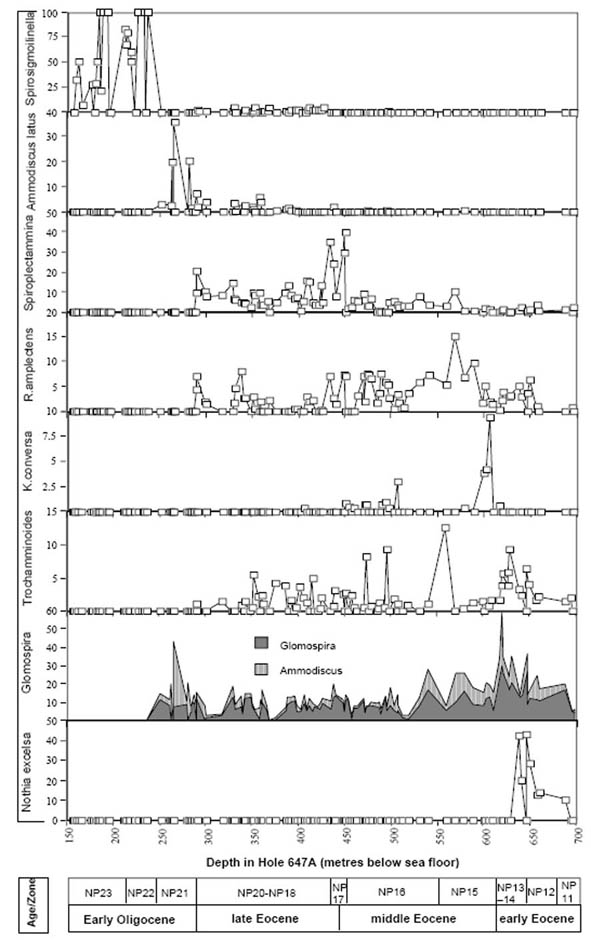
Figure 45. Relative abundance of selected species in the Eocene to Lower Oligocene of ODP Hole 647, southern Labrador Sea. Percentages are calculated as percent of agglutinated foraminifera (i.e., calcareous benthic species were removed from the matrix, to simulate a wholly agglutinated assemblage). Data are from Kaminski (1988).
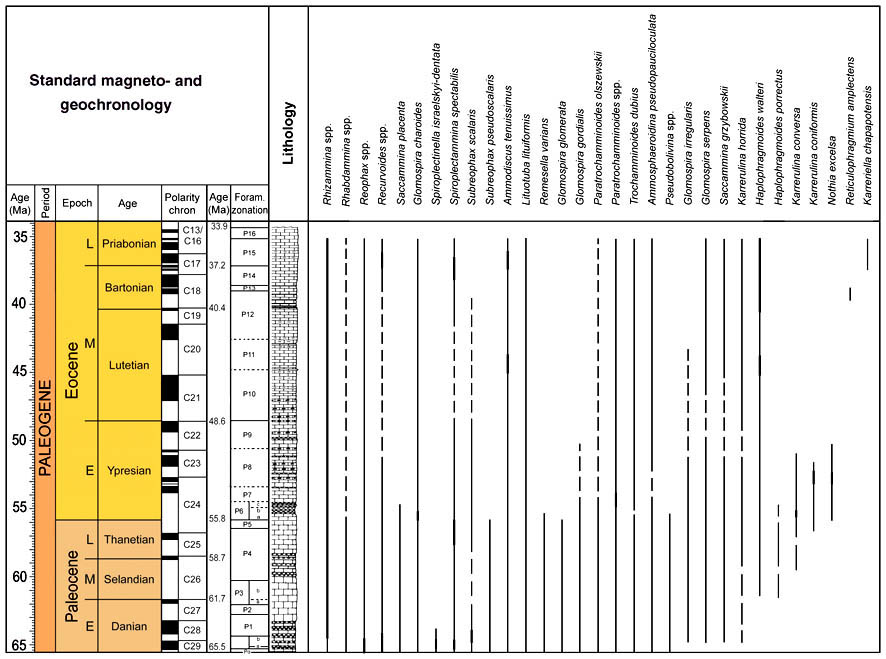
Nine DWAF acmes are recognised in the Eocene to lower Oligocene at Site 647 and at other Atlantic and Norwegian Sea sites (Sites 959, 985, 1047, 1048). These are: The "Strangelove Ocean" spiroplectamminid acme, The PETM Glomospira Acme, a lower Eocene N. excelsa acme, an early/middle Eocene glomospirid acme, a Karrerulina acme, a middle Eocene Reticulophragmium amplectens acme, a middle/late Eocene Spiroplectammina acme, a latest Eocene-early Oligocene Ammodiscus latus acme, and an early Oligocene Spirosigmoilinella acme. Some of these acmes can be correlated with similar events occurring at onshore
localities in the Western Tethys (northern Spain, Moroccan Rif, Italian Appenines, Western Carpathians). The main acmes are presented graphically in figure 46.
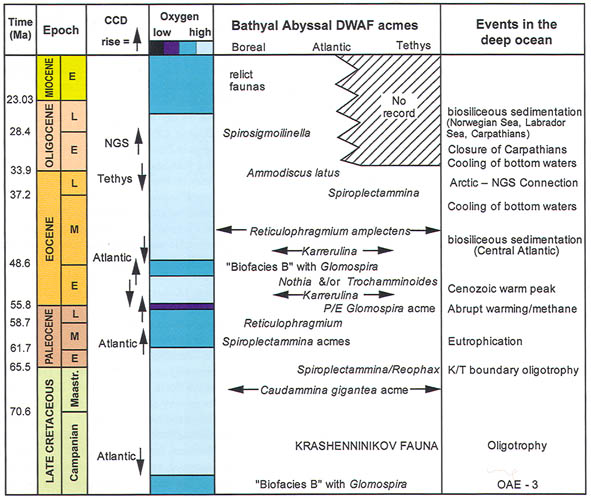
Figure 46. Summary of the areal extent of DWAF acmes, and palaeoceanographic parameters in the Atlantic-western Tethys region. Compiled from various sources and personal observations.
The "Strangelove Ocean" Spiroplectamminid Acme.
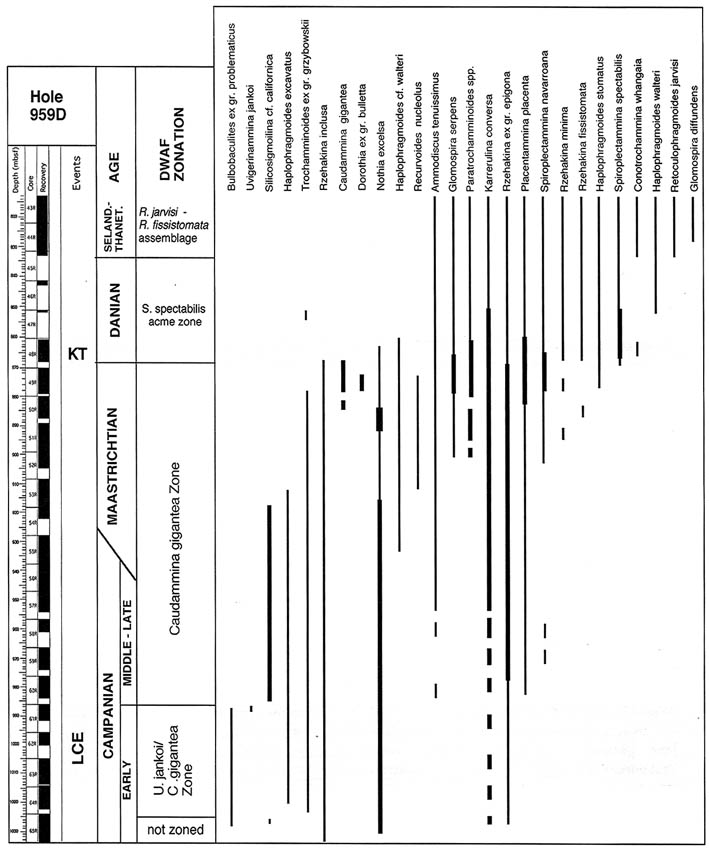
In the Sopelana section in northern Spain, the K/P boundary interval was studied by Kuhnt & Kaminski (1993). The foraminiferal assemblages within and directly above the boundary clay is dominated by infaunal agglutinated species such as Reophax, Subreophax, Ammobaculites and Recurvoides. Immediately above the boundary clay, a Spiroplectamminid-dominated biofacies characterises the top of Zone P0 and part of the P. eugubina Zone. This event has been subsequently found in the Caravaca section of Southern Spain (Coccioni & Galeotti, 1994), and we have observed it in the Contessa, Bottaccione, Petriccio, and Monte Conero sections of Central Italy. In the Italian sections, the acme is actually comprised mainly of Spiroplectinella israelskyi, but S. spectabilis is also present. The earliest Paleocene "Spiroplectammina event" (sensu Kuhnt & Kaminski, 1996) has now been observed at localities throughout the Atlantic and Mediterranean area, where the K/P boundary strata are preserved in deep-water settings. Peryt et al. (1997) found it in flysch deposits in Austria, and at ODP Site 959 in the eastern Atlantic, a peak in Spiroplectammina spectabilis was discovered by Kuhnt et al. (1998) in Cores 959D-48R-49R (see figure in taxonomic chapter). Most recently we have found this acme in the Indian Harbour M-52 well (Labrador Margin), housed in the BP Collection at the Natural History Museum, London.
Late Paleocene Reticulophragmium acme (Boreal Seas)
The evolution of alveolar structure among the organically-cemented deep-water agglutinated foraminifera took place concurrently along several lineages, in different oceanic basins during the mid to late Paleocene. Oceanic organic productivity as indicated by the ?13C ratios of pelagic carbonate was increasing steadily during the Paleocene (Corfield et al., 1991). Increased levels of sea-floor organic flux would have resulted in enhanced oxygen minimum zones below the thermocline and low values of oxygen saturation in the deep ocean. During the middle part of the Paleocene, the Atlantic Ocean became poorly oxygenated (Mountain et al., 1992), resulting in the deposition of dark coloured sediments in the deep basins. If the evolution of complex avleolar structure was a morphological adaptation to life in a poorly oxygenated environment, it is reasonable to assume that this evolutionary trait would evolve independently in different species. Several studies have pointed out the polyphyletic nature of the earliest Cenozoic cyclamminids (Berggren & Kaminski, 1990; Charnock & Jones, 1997), yet the group as a whole still suffers from a lack of detailed morphometric studies that would serve to document and understand the nature of its evolution. The single fact that is becoming increasingly well established is the observation that different species have evolved in different marginal basins of the North Atlantic during the Paleocene (Kaminski & Crespo de Cabrera, 1999).
In the North Atlantic petroleum basins the proportions of "primitive Reticulophragmium" increased throughout the late Paleocene, culminating in an acme just below the Paleocene/Eocene boundary. The "Reticulophragmium pauperum Zone" in the North Sea, Norwegian Shelf, and Western Barents Sea encompasses this acme. Various species of "primitive Reticulophragmium" evolved independently from various Haplophragmoides ancestors, mostly during the mid- to late Paleocene. In the Atlantic sector, for example the Indian Harbour well, the dominant species is Reticulophragmium garcilassoi, whereas in the Norwegian Sea we find R. pauperum and a robust Reticulophragmium. The genus Reticulophragmoides first appeared in the mid-Paleocene in the Atlantic and Norwegian Sea. An interesting primitive form of Reticulophragmium with few alveoles is R. vidonioensis from the Vidoño Shale of eastern Venezuela (Kaminski & Crespo de Cabrera, 1999). Several seemingly endemic species such as Reticulophragmium arcticum and its relatives are observed in the Paleocene of the Alaskan offshore (Petracca, 1972), in the deltaic sediments of the Beaufort Sea (McNeil, 1997), and the Paleocene of Spitzbergen (Nagy et al. 2000).
The genus Reticulophragmium is common mainly in neritic to bathyal facies, along the Atantic margins, probably preferring more eutrophic conditions. Equally intriguing is the observation that primitive Reticulophragmium is completely absent in the more oligotrophic and well-oxygenated Scaglia Rossa of Central Italy, and the abyssal ODP holes in the Eastern Atlantic.
The PETM Glomospira Acme. Age: NP10
We use the term "Glomospira" in the broadest sense, as this distinctive biofacies may contain several species that are (or should be) placed in difference genera, including various species of Glomospira, Glomospirella, and Ammodiscus. A short-lived maximum in the proportions of the glomospirids is observed at the Paleocene/Eocene boundary at Gubbio (Galeotti et al. 2004). The glomospirid maximum at Gubbio is found within a "dissolution horizon" within the normally reddish marly limestone of the upper Scaglia Rossa Formation. Its position just above the Paleocene/Eocene boundary in Gubbio has been determined based on the carbon isotope record (Galeotti et al. 2004). Recent ODP drilling in the South Atlantic suggests the CCD shoaled to at least the level of the Oxygen Minimum Zone (J. Zachos, personal communication), probably in response to the release and subsequent oxidation of gas hydrates. In the Contessa Road section the acme occurs in the 5-m interval above the P/E boundary that corresponds exactly to calcareous nannofossil Zone CP8. The most abundant species is Glomospira charoides, which is often of very small dimensions.
The acme of Glomospira charoides seems to be of use for stratigraphic correlation across the P/E transition in deep-water settings. Although not accurately dated, this is probably the same event that is present over a 12-m interval in ODP Hole 900A, and over a 3-m interval in Hole 897C off Iberia. Although the base of the Eocene was not recovered at Site 1067, a maximum in Glomospira is found in Cores 1067A-11R and -12R, which is dated as early Eocene (Zones NP11-12). A remarkable bloom of G. charoides occurs in the Trabakua Pass section in the Basque basin, Spain, where it forms 50% of the assemblage in the lowermost Eocene (Coccioni et al., 1995; Orue-Extebarria et al., 1996). The Glomospira-Ammodiscus assemblage reported by Kaminski et al. (1996) from the Paleocene-Eocene Talaa Lakrah section in Northern Morocco might be coeval. In the Polish Carpathians, the event is widely distributed and it constitutes the basal Eocene zone in the DWAF zonation of Olszewska (1997).
The widespread occurrence of a diminuative Glomospira biofacies in the earliest Eocene can be interpreted in terms of ocean-wide oligotrophy, as also suggested by the calcareous nannofossil record (Bralower et al., 2002). In a study of modern benthic foraminifera along a productivity gradient in the Mediterranean, De Rijk et al. (1999) found that Glomospira charoides characterises the deepest, most oligotrophic sector of the Eastern Mediterranean. In Gubbio, the facies is preceded by a marked decrease in the relative abundance of deep-infaunal forms, suggesting a decrease in the carbon flux to the sea-floor (Galeotti et al., 2004). While the duration of the earliest Eocene dissolution facies caused by the release of methane hydrates is only on the order of 200,000 years (Dickens, 2000), we expect that the duration of the "PETM Glomospira acme" may vary from place to place according to the trophic conditions. It is conceivable that the event represents a longer period of time at deeper, more oligotrophic sites because of the more severe oligotrophy generally found at deep-sea localities.
Nothia excelsa acme. Age: NP11-NP13
In Hole 647A, the species N. excelsa comprises a maximum of 42% of the DWAF assemblage at a depth of 646.9 mbsf (Fig. 45). In the eastern Atlantic Kuhnt & Collins (1996) reported a maximum of this species in one sample dated as lower Eocene at Site 897 (Sample 897D-3,CC). It is also present in elevated numbers in the "lower Eocene" at Site 900, where it occurs in Core 900A-72, approximately 30 m above the "PETM Glomospira acme"; and in Hole 1068A in the same relative position (5-10 m above the Glomospira acme). We also observed high proportions of N. excelsa in samples from the lower Eocene of the Contessa Road section in Italy, over the interval corresponding exactly to Chron C24n, which correlates to Zone NN12.
The Early/Middle Eocene Glomospira Acme. Age: NP13-NP14
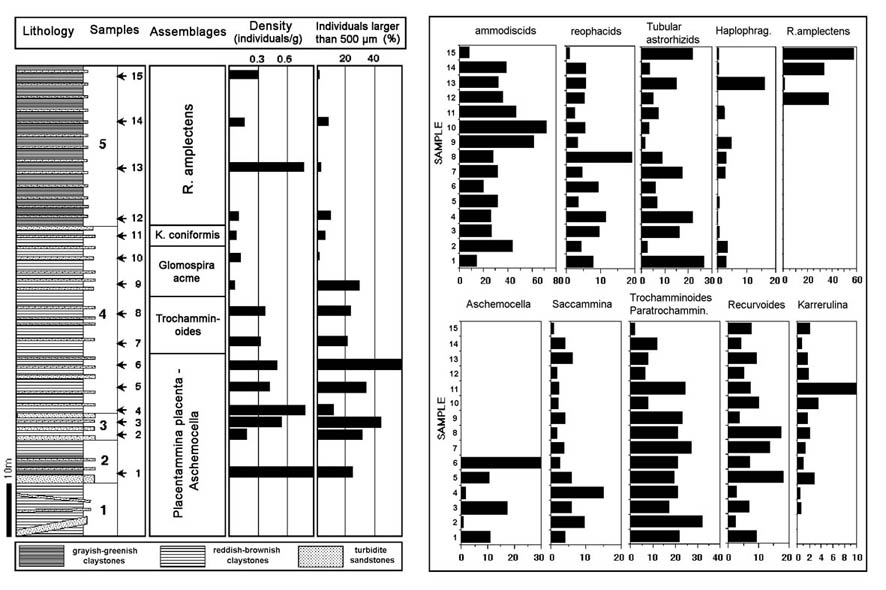
The occurrence of glomospirid-dominated biofacies is not a unique event. These events are well-known from the Cretaceous, where they have been termed "Biofacies-B" by Kuhnt & Kaminski (1989). In Hole 647, the Glomospira-Glomospira group comprises a maximum of 30% of the assemblage at 619.6 mbsf. In this hole, the occurrence of Ammodiscus seems to weakly covary with that of the glomospirids (r2=0.28), and this genus makes up 28% of the assemblage at this level. In the Labrador Sea, this E/M Eocene Glomospira assemblage is present within a 20-m thick dissolution horizon containing red and green variegated claystones. This is the only interval in the whole of the core that contains horizons of red clays. Backtracking indicates the CCD in the Labrador Sea had risen to a level above 2,500 m during the deposition of this "Glomospira facies".
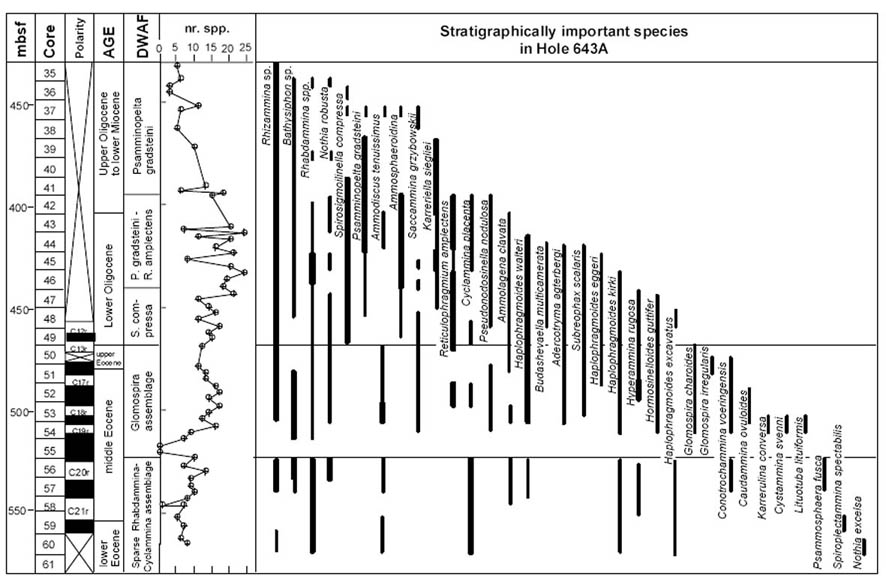
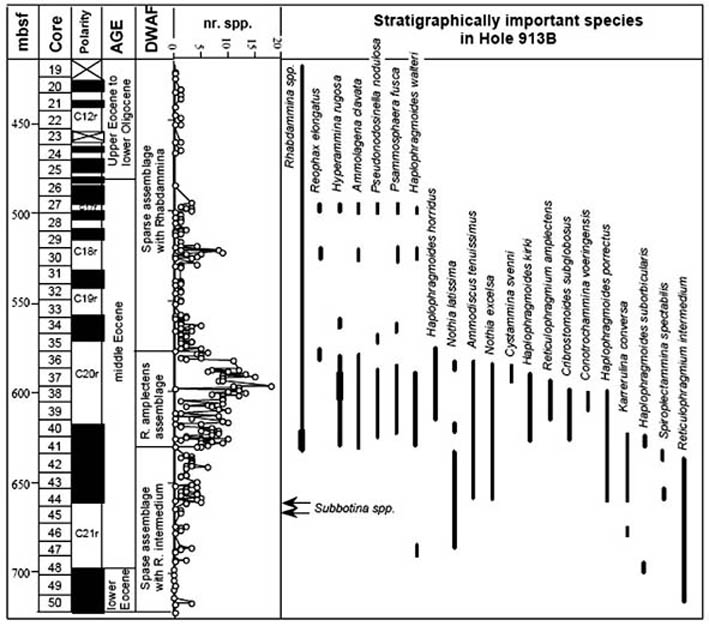
This second Glomospira acme is clearly present in the data from Hole 900A, where it is present as the most abundant non-tubular taxon in Sample 900A-70R-2, 57-59 cm (Kuhnt & Collins, 1996). This sample coincides with the FO of Reticulophragmium amplectens at this locality. Small numbers of Glomospira still occur at ODP Site 643 on the Vøring Plateau (Kaminski et al., 1990), but none were found at more northerly Site 913 in the East Greenland Basin (Osterman & Spiegler, 1996).
Karrerulina acme. Age: early NP15
A sharp peak in Karrerulina conversa is observed at the top of the "Glomospira assemblage" in Hole 647A. The species comprises 9.5% of the DWAF assemblage at 605.8 mbsf. At Site 900, the event is observed 1.5 metres above the second Glomospira acme, in the next higher sample (900A-70R-1, 69-72 cm). At Site 1067, Karrerulina conversa and Karrerulina coniformis together comprise an acme in Cores 1067A-5R and -6R. The acme begins at the base of Zone NN15 (subzone CP13a) and persists to the lower part of subzone CP13b).
This Karrerulina event is curious, as we have also observed an acme of K. conversa in the upper part of the lower Eocene in the North Sea (Gradstein et al., 1994), and it is observed as far north as ODP Site 913 in the East Greenland Basin (see: Table 2 of Osterman & Spiegler, 1996). It occurs in Morocco (Kaminski et al., 1996) in more or less the same relative position. However, the latter areas the chronology of the event is not as well constrained.
In Moravia, where planktonic foraminifera have been reported from the Carpathian deposits, Hanzlíková (1983) noted an acme of Karrerulina within the M. formosa-M. aragonensis Zones (undifferentiated) in the Magura Basin. Most recently, Bąk (2004) reported an acme of Karrerulina in the lower Eocene of the Dukla basin. We have also observed elevated proportions of Karrerulina conversa in Lower Eocene variegated claystones from the Silesian Unit near Biecz. In the Contessa Section in Italy, we found a maximum in K. conversa at 38 m above the C/P boundary within nannofossil Zone CP9, and another Karrerulina spp. acme at 100 m in Zone CP13. As this is the case, there may be more than one Karrerulina acme in other areas as well. It may be important to differentiate between the lower Eocene K. conversa acme and the middle Eocene mixed Karrerulina acme. The stratigraphic occurrence of the various Karrerulina species may be controlled by palaeoceanographic factors such as changes in organic matter flux, since modern representatives of this group appear to thrive infaunally in the abyssal clays of the North Atlantic (Kuhnt et al., 2000).
Reticulophragmium amplectens acme. Age: NP15 to early NP16.
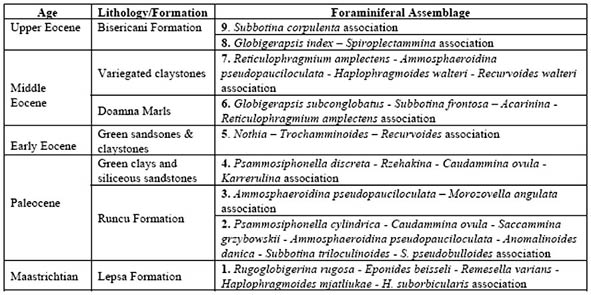
The species Reticulophragmium amplectens was one of the taxa originally used by Grzybowski (1898) in his zonation of the Potok Oil Field, near Krosno, Poland. The species is very widely distributed throughout the Western Tethys and North Atlantic.
In Poland and the North Sea, R. amplectens tends to be most abundant in the early part of its range. This trend is confirmed at Site 647, although the relative abundance of this species is much lower in the abyssal Labrador Sea than it is on the continental margin, where it can dominate the assemblage. A broad maximum in R. amplectens is observed between 588.4 and 518.8 mbsf in Hole 647A, with a maximum abundance of 15% recorded at 569.1 mbsf. At this site, its FO is observed in the lower part of Lower Eocene Zone NP11, and its LO is exactly at the level of the Eocene/Oligocene boundary as determined by calcareous nannofossil biostratigraphy. At Site 900 on the Iberian Abyssal Plain, the R. amplectens acme was observed 10 m above the Karrerulina acme, in Core 900A-69R-1. At this level it was the dominant species (Kuhnt & Collins, 1996). At Site 1067, it begins in Core 1067A-5R, immediately above the Karrerulina acme (Kuhnt & Urquhart, 2001).
Reticulophragmium amplectens is one of the most widely known species described from the Carpathian flysch, and has been reported from Poland, Slovakia, Moravia, the Ukraine, and Romania by numerous authors. Reticulophragmium amplectens figures prominently in most zonal schemes using agglutinated foraminifera, both within the Carpathians and in other regions. In Poland, its first occurrence was reported in the Early Eocene (Zone NP12) by Olszewska & Smagowicz (1977). Its total range in the Polish Carpathians was given as Early to Late Eocene by Morgiel & Olszewska (1981), and its partial range and peak occurrence characterises the Middle Eocene Cyclammina amplectens Zone of Geroch & Nowak (1984). In Moravia, the first appearance of abundant R. amplectens was reported by Hanzlíková (1983) from Zone P10 based on the occurrence of Acarinina densa, A. pentacamerata, and M. aragonensis. In the central North Sea, King (1989) used the LCO (last consistent occurrence) of R. amplectens to define the upper limit of his Zone NSA4 (R. amplectens Zone), which he regarded as Early to Middle Eocene. The middle Eocene R. amplectens acme is also found at the deep Norwegian Sea Sites 643 and 913 (Kaminski et al., 1990; Osterman & Spiegler, 1996), and it characterises the top of the Torsk Formation in the Western Barents Sea (Nagy et al., 1997).
Spiroplectammina acme. Age: NP17
The species Spiroplectammina spectabilis is present throughout the Eocene at Site 647, but its relative abundance increases abruptly to 40% of the assemblage at 450.1 m. The abundance acme continues to 434.4 m, where its abundance is still 34%. Higher in the Hole 647A, its abundance is lower, but it still reaches 15% in some samples. Its LO correlates exactly with the level of the Eocene/Oligocene boundary in Hole 647A. The "Spiroplectammina acme" in Hole 647A also includes the somewhat larger species S. trinitatensis (Cushman & Renz, 1946), which co-occurs with S. spectabilis in the younger (upper Eocene) part of the section. This event is apparently coeval with the Spiroplectammina spectabilis acme observed within Zone CP15a at 142.5 to 155 m in the Contessa Section in Italy.
The stratigraphic significance Spiroplectammina spectabilis is not unequivocal, because as is the case with Glomospira, the occurrence of this species seems to be linked with the trophic continuum. Spiroplectammina is absent from the highly oligotrophic abyssal sites in the eastern Atlantic. Kuhnt & Collins (1996) did not record a single specimen of this species from the Iberian ODP sites, and Kuhnt & Urquhart (2001) reported rare specimens only from the Paleocene at Site 1068. It was also absent from our samples from the Paleocene-Eocene Talaa Lakrah section in Northern Morocco. However it can be very common at times, and forms distinct acmes in the Paleocene and Eocene at Gubbio. It can also be quite common in middle-upper Eocene samples from the Carpathian flysch. Bratu (1975) recorded an assemblage in lowermost part of the upper Eocene Bisericani Formation in the Romanian Carpathians. In the Atlantic, Spiroplectammina spectabilis is better known to form acmes in the Paleocene rather than in the Eocene. It is known as a "disaster species", occuring just above the Cretaceous/ Paleogene boundary at ODP Site 959 in the equatorial Atlantic (Kuhnt et al., 1998) and at Gubbio, Italy (Kuhnt & Kaminski, 1996). In the Barents Sea, its maximum occurrence is observed in the middle part of the Paleocene, while in the Central North Sea and in the Contessa section it is common in the upper Paleocene. Its occurrence in the deep-sea sites is most likely linked to an increase in sea-floor carbon flux. At Site 647 the biosiliceous flux increases uphole throughout the upper Eocene (Bohrmann, 1989), suggesting increasingly more eutrophic conditions.
Ammodiscus latus acme. Age: latest NP20-NP21
The species Ammodiscus latus occurs in low numbers throughout the middle and upper Eocene at Site 647, but its relative abundance increases rapidly at 281.2 mbsf. This is just below the Eocene/Oligocene boundary as determined by the age model for Hole 647A, which places the boundary at ca. 290 mbsf within Core 647A-31R (Baldauf et al., 1989). Ammodiscus latus can be very common and reach very large dimensions in the lower Oligocene along the Labrador Margin - Northern Grand Banks. In the Carpathians, the Ammodiscus latus Partial Range Zone was defined as the interval from the FO of the nominate taxon to the FO of Reticulophragmium rotundidorsatum (Geroch & Nowak, 1984). Ammodiscus latus can occur commonly in upper middle Eocene to upper Eocene strata in the Carpathian flysch basins. The species is also known from other localities in the North Atlantic (Trinidad, North Sea).
Spirosigmoilinella/Psamminopelta acme (Boreal Seas). Age: NP22-23.
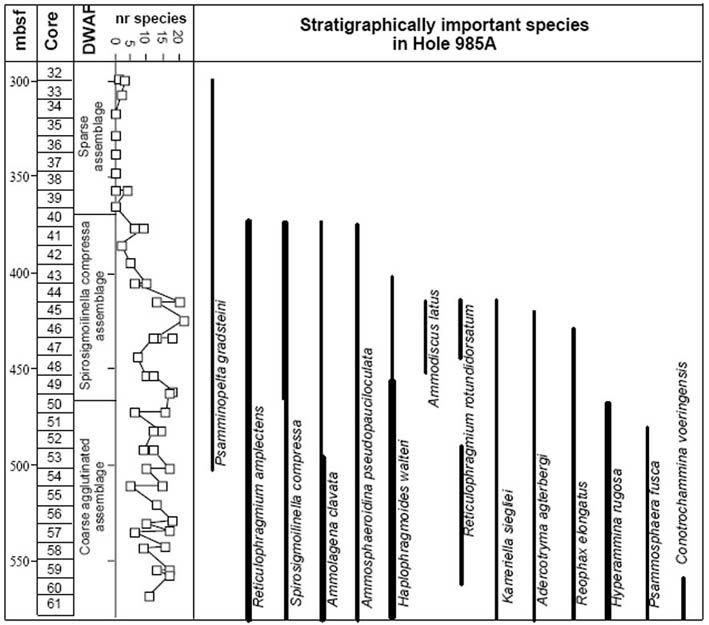
The diversity of agglutinated foraminifera declines abruptly across the Eocene/Oligocene boundary in Hole 647A. The Lower Oligocene at this site contains a depauperate DWAF assemblage, in sediments that are increasingly biosiliceous. At 237.1 mbsf the residual assemblage is comprised only of the species Spirosigmoilinella compressa and Psamminopelta gradsteini. A similar faunal change is known from the Oligocene of DSDP/ODP holes in the Central Norwegian Sea (Verdenius & van Hinte, 1983; Kaminski & Austin, 1999). This assemblage appears to be restricted to the northern North Atlantic.
Back on top | Introduction | North Atlantic | Beaufort-Mackenzie, Arctic | Western North Atlantic | Eastern Atlantic ODP | Western Tethyan Flysch | Pelagics | Summary