 |
|
|
|
updated to follow: Stratigraphic Guide to the Cromer Knoll, Shetland and Chalk Groups of the North Sea and Norwegian Sea. Felix M. Gradstein & Colin C. Waters (editors), Mike Charnock, Dirk Munsterman, Michelle Hollerbach, Harald Brunstad, Øyvind Hammer & Luis Vergara (contributors). Newsletter on Stratigraphy, vol 49/1 pp71-280, 2016
Foraminiferal biozonation | Dinocyst biozonation
Two practical Cretaceous biostratigraphic zonations are outlined for the petroleum sectors of the northern North Sea and Norwegian Sea, one using mainly Foraminifera and the other dinoflagellate cysts.
The quantitative foraminiferal zonation updates Gradstein et al. (1999). It combines shelly and some organic walled fossil events in one interval zonation, using mostly last occurrence and last common occurrences of over 100 taxa in over 30 wells. The taxa listed are directly from the samples in the Norwegian Sea and North Sea wells on which the zonation is based.
The separate dinoflagellate cyst zonation is a literature zonation applicable to the North Sea and Norwegian Sea. The taxa and zonation are based on available peer-reviewed palynological publications, and completed with unpublished PhD theses and collected contributions from course manuals. Note that the dinoflagellate zonation incorporates more recent literature than was used when the quantitative micropaleontology/palynology zonation was calculated; hence taxonomic details differ between both zonations but are a minor issue.
Modern biostratigraphy, particularly when well data are involved, emphasizes fossil events over fossil ranges along a depth or time scale. A biostratigraphic event is the presence of a taxon in its time context, derived from its occurrence in a rock sequence. Such events are unique and irreversible. Event positions are preferred levels (like last occurrence or last common occurrence) along the stratigraphic range of taxa. The stratigraphic ranges of taxa often change from basin to basin, with concommittant changes in event positions. The relative and/or linear positions of these events are influenced by many factors, paleoecologic, evolutionary, paleobiological and taxonomic in nature. Subjective judgement of correlations plays an important role also. Events may change relative position from region to region, or even across any one region and also between schemes by different authors. It may not always be easy to point to clear reasons for such differences. Below, we will try to outline differences between zonal event schemes, using Foraminifera.
The biostratigraphy of Lower and Upper Cretaceous foraminifers in NW Europe has been studied by a number of authors since the 1960's. A brief review of biostratigraphic schemes developed for the NW European siliceous mudstone facies is useful in order to understand the microfossil records from the eastern sedimentary wedge of the Norwegian-Greenland Sea.
Northwest Germany - The work from H. Bartenstein and co-workers on the benthic foraminifers of the Saxon Basin has lead to the publication of a detailed biostratigraphic scheme for NW Germany (Bartenstein & Bettenstaedt, 1962). This was followed by a proposal for a general biostratigraphic zonation for the Lower Cretaceous using more cosmopolitan taxa (Bartenstein, 1978). The biostratigraphic zonation of the NW German Lower Cretaceous was based on the stratigraphic succession of 75 foraminifer, and 12 ostracod species in middle Valanginian to lowermost Cenomanian of Saxony. The stratigraphic ranges of benthic foraminifers were calibrated to the standard ammonite zones. The zonal scheme mainly uses calcareous benthic foraminiferal species more typical of shallow marine sediments, such as Citharina, Epistomina, and ornamented Lenticulina. Unfortunately, neither of these genera is common in the Cretaceous Norwegian Sea. Fewer than 15 of the Northwest German index species were commonly encountered in this study. Of these, the 11 taxa that do occur consistently offshore Norway, and are thus used for chronostratigraphic correlation, are listed in Table 2.1
Table 2.1

Northern Tethys Margin - Weidich (1990) published a zonal scheme for the Tithonian to Cenomanian based on his studies of the Northern Calcareous Alps in Bavaria and Austria. His scheme divides the Lower Cretaceous into 10 zones, based on first occurrences or acmes of nominate zonal taxa. Weidich's zonation is based primarily on calcareous benthic foraminifers, but because his studied sections were deposited in deep water, there is a greater possibility for correlation with the Norwegian Sea region. Several of the index taxa are also found in the North Sea, which verifies Weidich's selection of taxa that are largely cosmopolitan. However, Weidich's zonal scheme was only tentatively correlated to the standard chronostratigraphy, and calibration of Weidich's samples by means of planktonic microfossils is needed to refine the age of his zones. Table 2.2 gives a list of stratigraphically important cosmopolitan species from Weidich's study, which also occur in the Norwegian Sea assemblages.
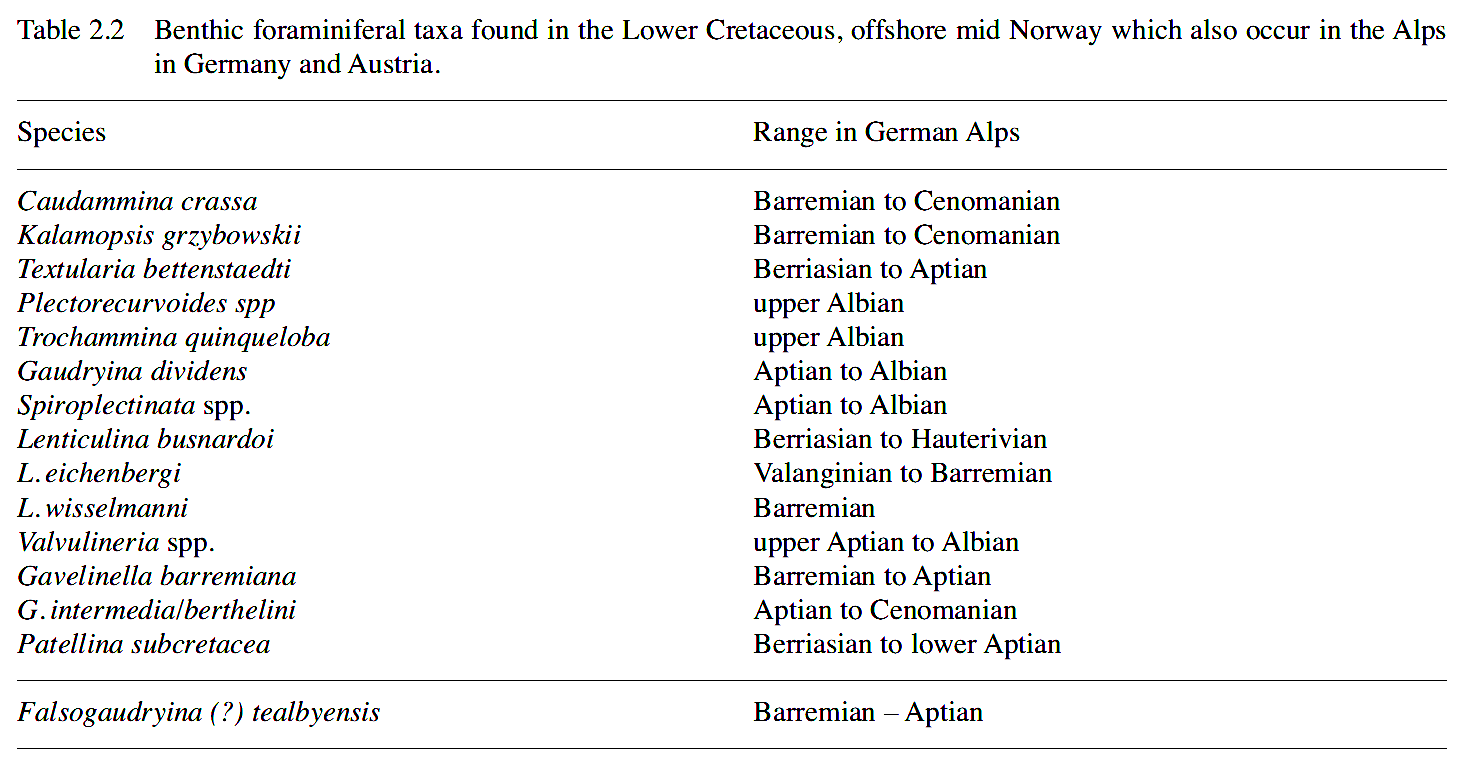
The biozonation of Geroch and Nowak (1984), with additions by Geroch and Koszarski (1988), provide a reference frame for cosmopolitan deep-water agglutinated foraminiferal assemblages. The Lower Cretaceous portion of their scheme consists of 19 species in 7 zones, and the Upper Cretaceous of 17 species in 4 zones. All zones are based on the first occurrence of index taxa - so they are not equivalent to zones of the same name recognised in other areas, and are not directly applicable to cuttings samples in wells. Although these zones were originally developed for the Carpathian flysch deposits, parts of the Geroch and Nowak scheme apply to other deeper marine continental margins with Cretaceous 'flysch-type assemblages, as far away as the NW Australian margin (Kaminski et al. 1992), and the Pacific (Riegraf & Luterbacher, 1989).
Several of the nominal index species from this zonal scheme we have now observed also in the Norwegian seaway, including Verneuilinoides neocomiensis, Pseudobolivina variabilis, Pseudonodosinella troyeri (= Reophax minutus of Geroch & Nowak), Haplophragmoides nonioninoides, and Bulbobaculites problematicus. Other species we observed from Geroch & Nowak's (1984) range chart include Ammobaculoides carpathicus, Trochammina quinqueloba, Trochammina abrupta, Thalmannamina neocomiensis, Caudammina crassa, Hippocrepina depressa, Kalamopsis grzybowskii, Gaudryina filiformis ( = ? Gerochammina lenis), Caudammina ovulum, Rzekina epigona, Tritaxia dubia (= T. subparisiensis), Marsonella crassa and Dorothia oxycona. Many of these species were not previously reported from the Norwegian and North Seas.
Central North Sea - A foraminiferal zonal schemes for the Lower and Upper Cretaceous of the Central North Sea, developed by C. King and co-workers at the Paleoservices consulting company was published in the 'Stratigraphical Atlas of Fossil Foraminifera' (Jenkins & Murray, 1989); it links to the detailed study by Koch (1977) for the calcareous Upper Cretaceous of NW Germany. King et al. (1989) proposed two parallel zonations for the North Sea, subdividing the Lower Cretaceous into 12 zones and subzones, and the Upper Cretaceous in 11 zones and subzones (referred to in Figure 2.6). The "FCS" zonal scheme has applications in the shelf (chalk) facies of the southern sector of the North Sea, whereas the "FCN" zones are based on deeper-water taxa and can be used for the bathyal shale facies in the northern and central sectors. The FCN zonation scheme is most applicable to our study area, and the zonation offshore Norway resembles this scheme (referred to in Figure 2.6, and discussion below). The index taxa are widespread and common, which makes the scheme quite robust. However, little attention was paid by King et al. (op. cit.) to deep water agglutinated foraminifers which become more important northward. Below, we will discuss in detail similarities and differences of our NCF zonation to the FCN zonation.
Millennium Atlas - An important document for North Sea petroleum stratigraphy is the Millennium Atlas (Evans et al., 2003). For the Lower Cretaceous section of this excellent compilation, Copestake et al. (2003) created a sequence stratigraphic model with multiple biostratigraphic event linkage (albeit no zonation). No such overview was provided for the Upper Cretaceous in the Millennium Atlas, but the study by Bailey et al. (2009) provides a Chalk foraminifer benthic zonation.
Figure 2.4 shows the fossil events framework of Copestake et al. (2003) for the North Sea with linkage to the Hardenbol et al., (1998) ammonite biozones, magnetostratigraphy and Lower Cretaceous stages. Unfortunately, none of the Lower Cretaceous stages have yet been formally defined. In the zonal text below reference will be made to linkages with the Copestake scheme.
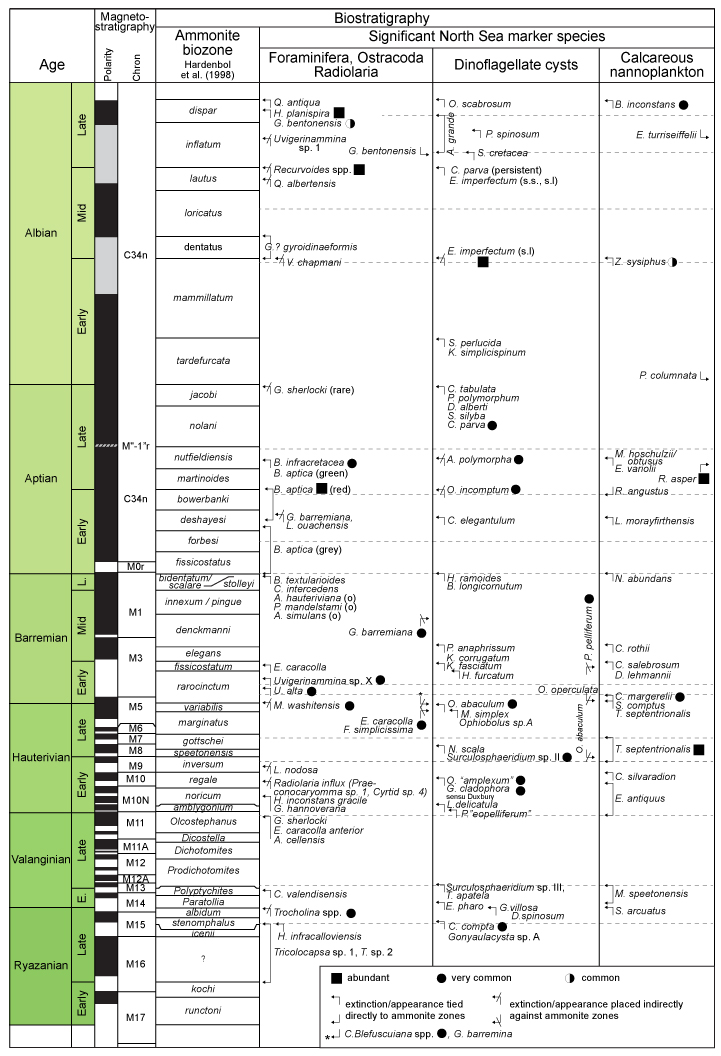
Our Cretaceous biostratigraphy for the North Sea and petroleum sector of the Norwegian Sea, offshore mid Norway spans almost 20 degrees latitude between 52 and 67 N. The transition from chalk and marls to siliciclastics takes place between 58 and 60 N, and modifies the biota content such that planktonic foraminifers become taxonomically much less diverse, and only Hedbergella taxa start dominating some levels.
The Cretaceous sedimentary succession may reach several kilometers in thickness, and basinward is buried under a Cenozoic ‘mudstone’ blanket’ of 2-3km. It may be subdivided in several broad units, listed below with generalized bio-and lithostratigraphy.
(1) Thin, multicoloured, marly sediments of late Hauterivian through early Barremian age, becoming dark coloured upwards, where the paleoenvironment changed from oxic to dysaerobic. These sediments herald the broad and global transgression in Cretaceous time over continental margins and other ‘highs’. The foraminiferal microfossil assemblage contains common Falsogaudryinella and nodosariids; ostracods also are common. It is a shallow marine, normal salinity microfossil association. The sediments belong in the Lyr Formation of the Cromer Knoll Group.
(2) Dark mudstones and minor sands, Aptian through early Cenomanian in age, with deeper water agglutinated foraminifers and monotypic (planktonic foraminfer) Hedbergella floods. The sediments largely belong in the Langebarn Formation of the Cromer Knoll Group, and were laid down in an upper bathyal environment, with a dysaerobic deeper watermass.
(3) Thick mudstone facies, with thin, slope-apron gravity-flow sands, with a locally impoverished benthic foraminiferal assemblage, deposited in an upper bathyal, oxic-dysaerobic facies, of late Cenomanian – Coniacian age. The sand units are classified in several lower-rank members of the Blålange Formation, including the well-known Lysing Member. Where Turonian sedimentation rates are low, a relatively rich planktonic foraminiferal assemblage may be found with Whiteinella, Hedbergella, Preaglobotruncana, Dicarinella and Marginotruncana. The AOE2 – Bonarelli (Plenus Marl) event at the Cenomanian –Turonian boundary is expressed in well logs with γ spike readings.
(4) Grayish, laminated mudstones, Santonian through Campanian in age, with locally thick sands in the distal Voering Basin have a mostly low-diversity deeper marine foraminifer assemblage, an Inoceramus peak, and common radiolarian/diatom taxa. The sediments belong in the Kvitnos and Nise Formations of the Shetland Group.
(5) Marly and sandy sediments of Maastrichtian age, with a low-diversity planktonic and benthic foraminiferal assemblage. Locally, and particularly southward, planktonics may occur in floods with monotypic Rugoglobigerina and Globigerinelloides. Sediments are grouped in the Springar Formation of the Shetland Group.
Organic-walled microfossil biozonations for the North Sea and Norwegian – Greenland Sea may be browsed, downloaded and printed from the Norlex website under www.nhm2.uio.no/norlex. Consultants frequently use proprietary zonations that borrow events from the literature. Relevant, non-proprietary charts of dinoflagelate-spore/pollen events may be found on the Norlex website under the heading ‘Biostratigraphy’.
Plate 2.1 illustrates three micro fossil taxa that are useful index taxa, and possibly are endemic to the greater North Sea region: Uigerinammina una, Ammoanita globorotaliaeformis, and Fenestrella bellii. The two first ones are restricted to the Uvigerinamina una Zone, late Middle to early Late Albian. The latter defines the Fenestrella bellii (total range) Zone, Early Campanian, and is an important marker in the deep Voering Basin (e.g. in the Gjallar well, 6704/12-1). Appendix 1 re-describes these taxa.
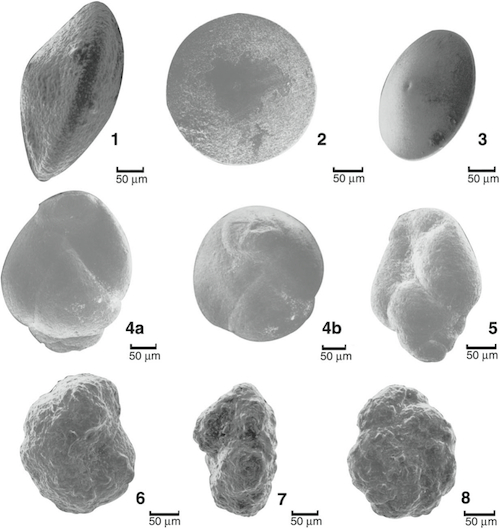 |
Plate 2.1. 1-3. Fenestrella bellii Gradstein andВ Kaminski. Well 6507/6-2, FenestrellaВ bellii zone, cts. 2320m, offshore mid-Norway; holotype in Fig. 1. Specimens were pyritized during burial. 4a ,b. holotype. Uvigerinammina una Gradstein and Kaminski. Uvigerinammina una zone, well 6507/6-2 at cts. 3110 m, offshore mid-Norway. The specimen in Figure 5 is from cts. 3040 m in the same well. В 6-8. holotype. Ammoanita globorotaliaeformis Gradstein and Kaminski. Uvigerinammina una zone, well 6507/6-2 at cts. 3110 m, offshore mid-Norway. |
The Cretaceous biozonation documented here uses foraminiferal taxa, dinoflagelates and some miscellaneous microfossils. Rather than trying to create a zonation that maximizes stratigraphic resolution, and often is difficult to apply over a broad region, we prefer to outline reliable zonal units that are easy to correlate. Hence, use was made of the Ranking and Scaling (RASC) method (Gradstein et al., 1985) to rank and scale different microfossil groups together. The lower Cretaceous, pre-Albian section is too fragmented, and well coverage insufficient to effectively apply event scaling using quantitative stratigraphic methods; that part of the stratigraphic section is ranked and zoned subjectively. Below, we first will summarize the procedure (from Gradstein et al., 1985 and 1999), prior to a discussion of the successive Cretaceous zones NCF 1-21.
Initially, the data set for RASC comprised the multiple microfossil event record in 37 wells, mostly LO (Last Occurrence in a stratigraphic sense) or LCO (Last common Occurrence in a stratigraphic sense) events of 550 foraminifers, some siliceous microfossils and dinoflagellates, for a total of 1873 records.
A majority of well samples we analysed self for shelly microfossils, complemented with selected information from fossil listings in well completion reports. The organic-walled microfossil record for the wells was obtained from consultants and colleagues (ex- Saga Petroleum). After several intermediate zonations, using RASC and its normality testing functions of the event record, five erratic events, and seven wells with low sampling quality (partly due to turbo-drilling) were deleted from the data.
The remaining 31 wells harbour 519 fossil events, with 1755 records. Principal wells are 6610/3-1, 6610/3-1R, 6607/5-2, 6607/5-1, 6607/12-1, 6507/2-1, 6507/2-2, 6507/2-3, 6507/6-2, 6507/7-1, 6507/7-2, 6506/12-5, 6406/2-1, 36/1-2, 35/3-1, 35/3-2, 35/3-4, 35/3-5, 35/12-1, 35/11-2, 35/11-5, 35/9-1, 35/9-2, 35/10-1, 34/7-21a, and 33/9-15.
Table 2.3 provides a summary of data properties and RASC results. Number of events in the optimum sequence with SD < ave SD, deals with the number of events that have a standard deviations below the average for the optimum sequence; the fact that 44 out of 72 optimum sequence events have a lower than average standard deviation is good. Stratigraphic coverage is relatively good, even that, as usual, only 97 out of 519 events occur in 6 or more wells, and only 72 events occur in 7 or more wells. Cretaceous dinoflagellate cysts in particular show large taxonomic diversity, but limited traceability for many taxa.
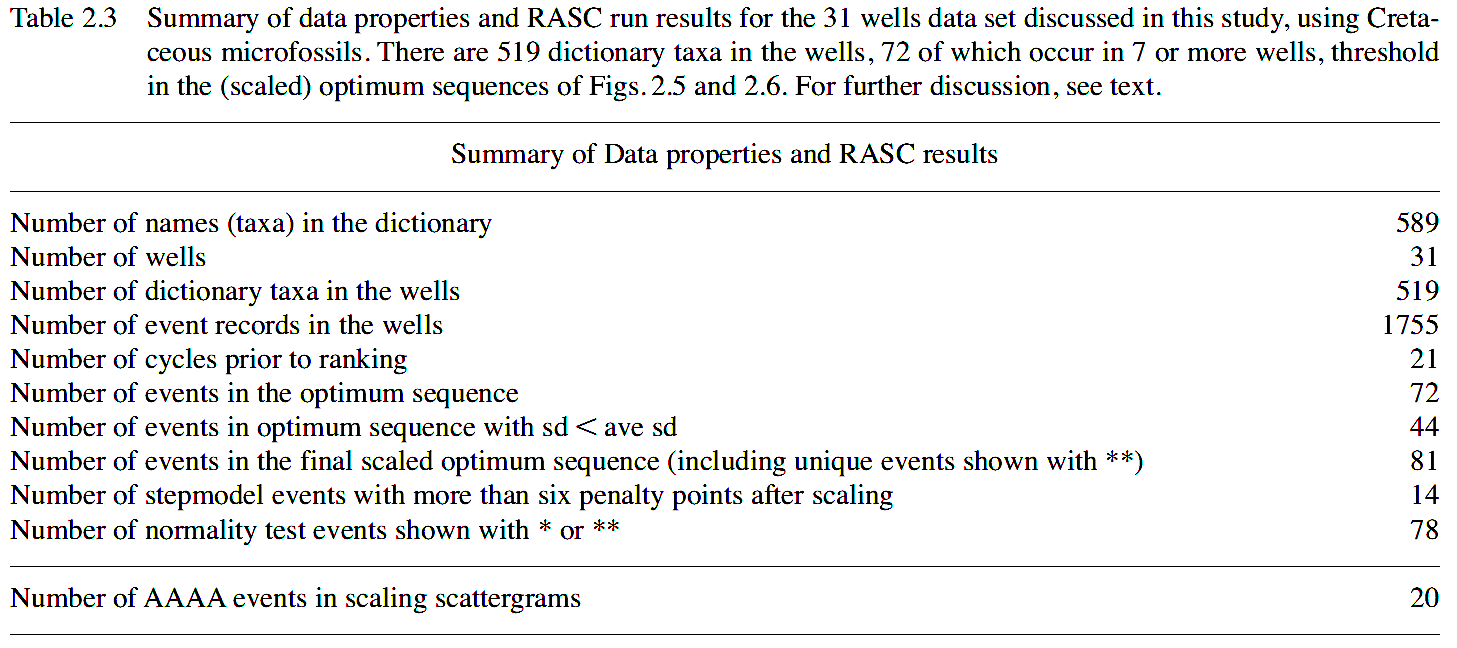
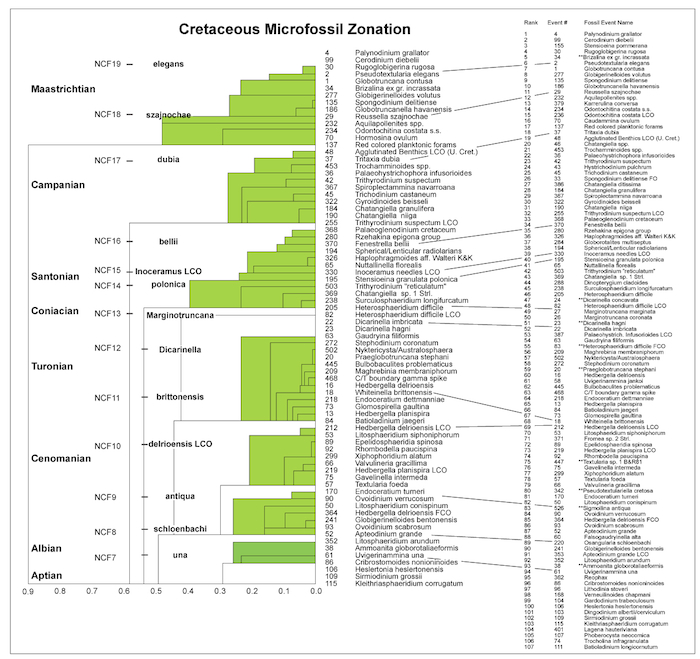
The Cretaceous optimum sequence (Figure 2.5, right) includes 98 events (‘tops’) that occur in 6 or more wells, spanning Hauterivian through Maastrichtian strata. Nine events, shown with double ** in front of the name, are socalled RASC unique events, meaning they have good calibration potential, but occur in fewer than threshold (6) wells. A simple RASC routine merges these unique events in the optimum sequence. ‘Range’ of taxa in the optimum sequence, a measure of the number of sequence positions an event spans (= rank positions in Figure 2.5, right), is always low, except for Fromea sp. 2 (5 rank positions), Dinopterigium cladoides (6 rank positions), Trithyrodinium 'reticulatum (5 rank positions), Gyroidinoides beisseli (5 rank positions), Chatangiella niiga (5 rank positions), and Chatangiella sp. (10 rank positions). The last occurrence record for these taxa is not reliable for detailed correlations in the wells examined. The optimum sequence serves as a guide to the stratigraphic order in which events in wells are expected to occur, hence it is both predictive, and serves in high-resolution correlation, as outlined below.
Figure 2.5 (left) shows the scaled optimum sequence for the Cretaceous in the 31 wells using events that occur in 7 or more wells. Nine events noted with ** occur in fewer than 7 wells, but are inserted as unique events for the purpose of zonal definition and chronostratigraphic calibration. The events listed in the optimum and scaled optimum sequences of Figure 2.5 represent average last occurrences (average ‘top), unless otherwise indicated; LCO stands for last common or last consistent occurrence, and FCO means first common or first consistent occurrence (in an uphole sense).
Over a dozen stratigraphically successive RASC interval zones stand out in Figure 2.5, middle Albian through Maastrichtian in age, which guide the discussion of interval, acme, and assemblage zones below. Resolution is poor in the lowermost and uppermost parts of the scaled sequence, and reflects lack of superpositional data, and condensation of ‘tops’ in the uppermost and lowermost Cretaceous part of wells.
 click to enlarge |
Figure 2.5 (right): Optimum sequence of Cretaceous events (foraminifera, dinoflagellates, miscellaneous microfossils and one physical log marker), offshore mid Norway, using the Ranking and Scaling (RASC) method on 1753 records in 31 wells. The arrow of time is upward. The majority of events are last occurrences (LO) in relative time; LCO stands for Last Common or Last Consistent Occurrence. Each event occurs in at least 6 out of 31 wells, leaving 97 events. The display also shows the relative stratigraphic position of nine unique events.В The optimum sequence is graphically tied to the scaled optimum sequence (= RASC zonation) to the left,В largely using the nominate zone markers. |
The scaled optimum sequence shows major breaks between several successive zones, including between the una and schloenbachi zones, delrioensis LCO and brittonensis zones, Marginotruncana and polonica zones, bellii and dubia zones, and particularly between dubia and szajnochae zones. The question what the meaning is of these large breaks, is best answered when we consider how RASC scaling operates. Scaling calculates the relative stratigraphic distance between successive events in the RASC optimum sequence. This relative distance is calculated from the frequency of cross-over of all possible pairs of events in the optimum sequence over all wells. Hence, large distances between successive events, corresponding to the larger RASC zonal breaks indicated above, signify strongly diminished cross-over between events that occur below and above the breaks. Reasons for these breaks thus should be thought in stratigraphic gaps, and paleoenvironmental changes. The reasons will be discussed in more detail below, but may be summarized as follows:
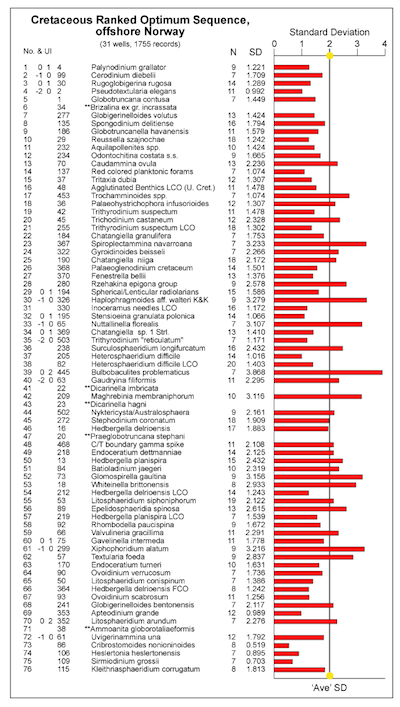
Figure 2.6 is an important zonal figure, it displays the RASC most likely order of events calculation using a threshold of 7 instead of 6 (right side of Figure 2.5). This means that each event used in the optimum sequence calculation must occur in a minimum of 7 wells, instead of 6 wells. To the left in Figure 2.6 is the optimum sequence of Cretaceous microfossil events in 31 wells, offshore mid-Norway. N is the number of wells sampled to calculate the s.d. per event (N > or = 7), and s.d. is the standardized deviation from the line of correlation in each well. The average standard deviation of 1.8903 is the sum of all s.d.’s, divided by the total number of events (=72) in the optimum sequence. The asterisk behind events indicates a s.d. that is smaller than the average s.d. From event s.d. theory it follows that events with a below average s.d. correlate the same relative stratigraphic level more faithfully from well to well than events with a higher s.d.
 click to enlarge |
Figure 2.6. Summary of event variance analysis results for the Cretaceous optimum sequence of Figure 2.5, but using a threshold of 7 instead of 6. This means that each event used for Ranking and Scaling (RASC) analysis must occur in a minimum of 7 wells, instead of 6 wells (as shown in Figure 2.5, right side). To the left in Figure 2.6 is the optimum sequence of Cretaceous microfossil events in 31 wells, offshore mid-Norway, where each event occurs in at least 7 out of 31 wells; N is the number of wells sampled to calculate the s.d. per event (N > or = 7), and s.d. is the standardized deviation from the line of correlation in each well. The average standard deviation of 1.8903 is the sum of all s.d.’s, divided by the total number of events (=72) in the optimum sequence. The asterisk behind events indicates a s.d. that is smaller than the average s.d. From event s.d. theory it follows that events with a below average s.d. correlate the same relative stratigraphic level more faithfully from well to well than events with a higher s.d. |
Within the major units, smaller zonal subdivisions may be recognized, using a combination of subjective judgement from individual well study, and objective subdivision from ranking and scaling analysis. Hence, the final zonation takes advantage of both subjective, and more objective analysis, and integrates the two into one framework for a nineteen fold NCF (North Sea Cretaceous MicroFossil) Zonation (Figures 2.7a,b). The numbering makes it easy to use for geologists and non-foraminifer biostratigraphers. As mentioned earlier, integration, using the RASC method, with dinoflagellate events, improves the applicability of both types of microfossils for regional biostratigraphy.
Besides the zonal numbers, the zones are also named after prominent foraminiferal taxa, which are easily recognized in the wells, and are relatively widespread. Some of the foraminiferal index taxa (like Trocholina, Blefusciuana, Globigerinelloides gyroidinaeformis, Hedbergella brittonenesis, Marginotruncana coronata and Dicarinella concavata) are rare due to the high-latitude setting of the study region, or poor preservation of carbonate tests in the siliceous mudstones.
Chronostratigraphic assignments are tentative, hampered among others by the absence, or scarcity of keeled planktonic foraminifers, including Rotalipora andВ the majority of Dicarinella, Marginotruncana, and Globotruncana taxa. Zones of relative short stratigraphic duration, like the Sigmoilina antiqua Zone, assigned to lower Cenomanian may be difficult to observe in ditch cuttings, particularly when the lithology is sandy. Zone definitions are bound to be refined with additional data, and further assessment of taxonomy, for example in the condensed Hauterivan - Barremian, and hiatus-riddled Aptian-middle Albian sections. Another zonal challenge is the thick, siliciclast interval in the inner Voering Basin, assigned to Campanian. The latter represents a poorly sampled section, probably laid down on a deeper marine, basin floor.
Stratigraphic range charts of principal taxa in the zonation of Figures 2.7a, b are in Figure 2.7a, b, c and in Figure 2.8. Taxon names followed by FO or FCO are events that help to define the lower level of the zones; similarly, taxon names followed by LCO or LO are events that assist with deliniation of the upper level of zones.
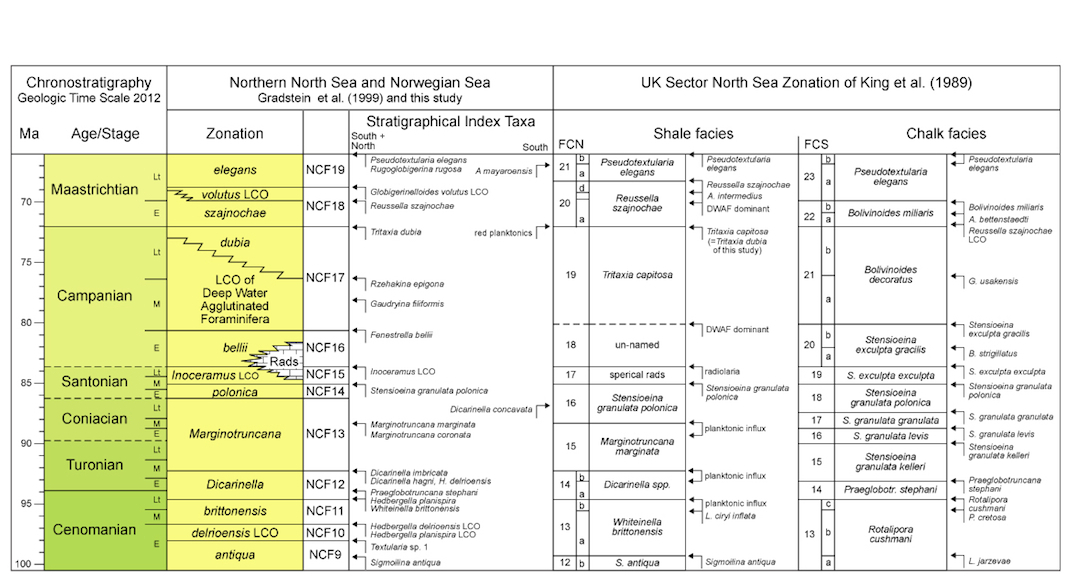 |
| Fig. 2.7b: Upper Cretaceous zonation for the Norwegian Sea, offshore mid-Norway, with stratigraphical index taxa for the region under study and taxa observed in its southern part only. The zonation is compared to the RASC model zonation using foraminifera and dinoflagellates. The zonation compares well with the North Sea zonation for shaly facies by King et al (1989), and is slightly more detailed in part. As expected, the chalk facies zonation of the North Sea does not compare or apply. |
| Click to enlarge |
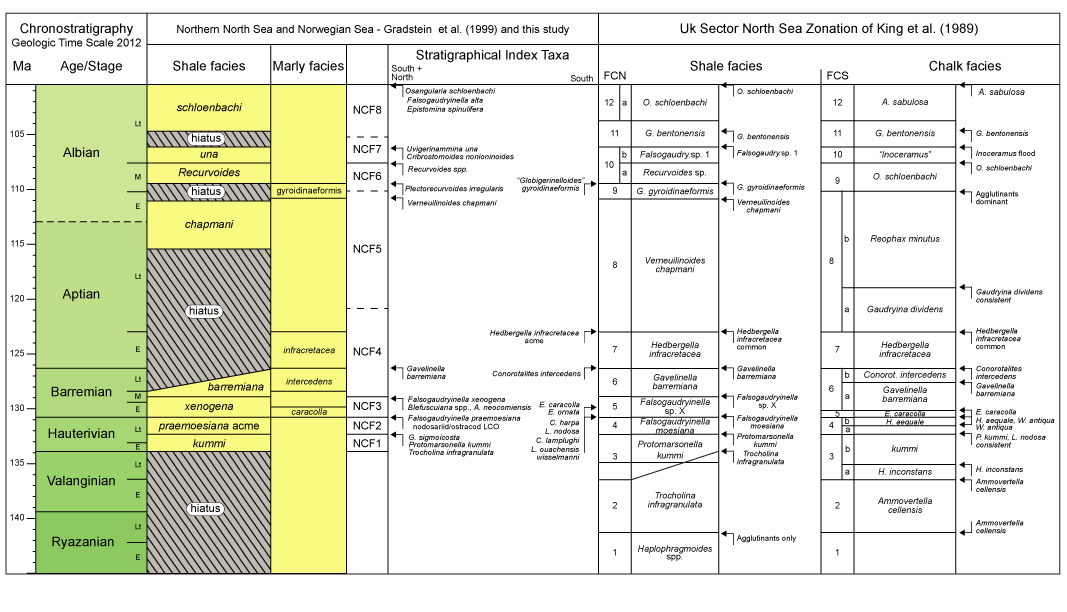 |
| Fig. 2.7a: Lower Cretaceous zonation, offshore mid- Norway with stratigraphical index taxa for shaly facies (more predominant northward), and for marly facies (more predominant southward). The zonation compares with the North Sea zonation for shaly (north) and for marly (south) facies of King et al. (1989), with which it shown taxa and zones in common. Seemingly, stratigraphic resolution decreases northward, but this is partly a bias from a more limited data set than available to King et al (op.cit.) in the North Sea. |
Stratigraphic ranges of Cretaceous foraminifera
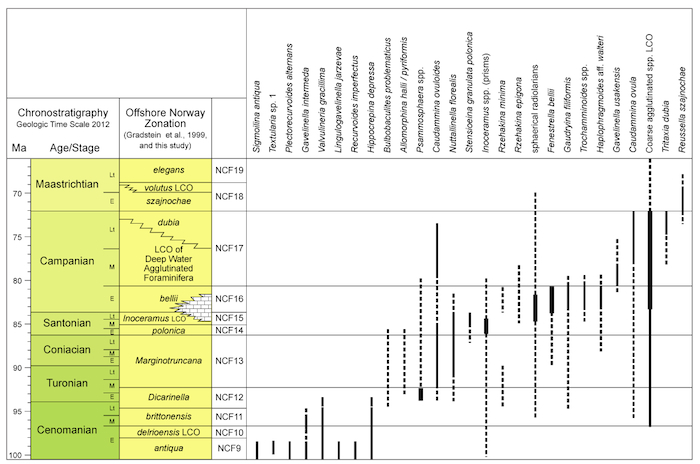
Figure 2.8c.В Stratigraphic В ranges of Upper Cretaceous benthic foraminifera, and miscellaneous index taxa, offshore mid-Norway, with the foraminiferal zonation established in this study. (Click to enlarge)
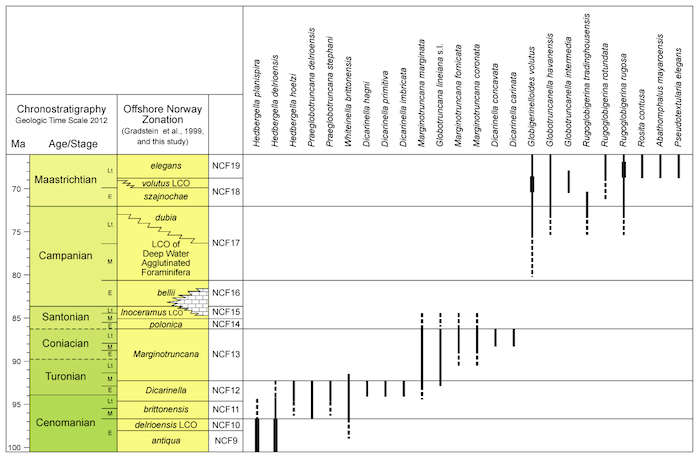
Figure 2.8b. Stratigraphic В ranges of Upper Cretaceous planktonic foraminifera index taxa, offshore mid-Norway, with the foraminiferal zonation established in this study. (Click to enlarge)
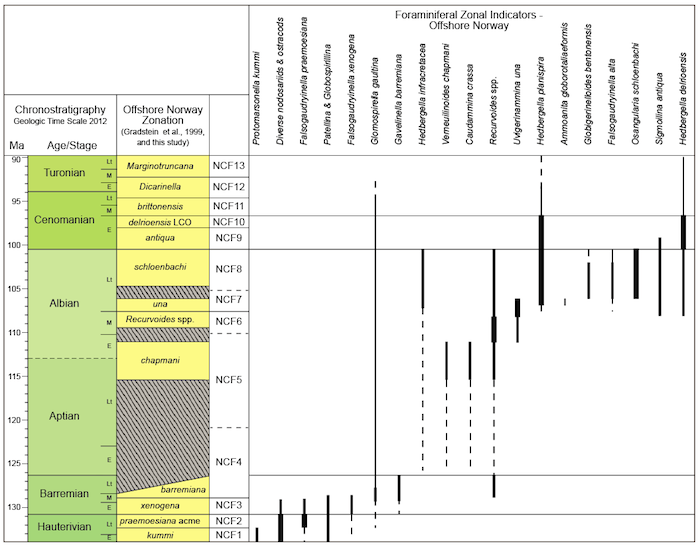
Figure 2.8a. Stratigraphic ranges of Lower Cretaceous index taxa, offshore mid-Norway, with the foraminiferal zonation established in this study. (Click to enlarge)
Protomarssonella kummi Zone = NCF1 Zone
This zone is defined on the upper range of Protomarsonella kummi, prior to the acme of Falsogaudryinella praemoesiana (Figure 2.7a). As described above, NCF stands for Norwegian Cretaceous (micro-) Fossil. The following taxa were observed in assemblages assigned to the zone:
Protomarssonella kummi LO
nodosariids and ostracods (common to abundant)
Falsogaudryinella praemoesiana FO (rare)
Patellina subcretacea
Trocholina infracretacea
Globospirillina neocomiana
Spirillina spp.
Lagena hauteriviana
Lenticulina aff. subalata
Lenticulina cf. busnardoi LO
Lenticulina munsteri (common)
Lenticulina heiermanni
Epistomina tenuicostata
Age: Hauterivian, probably early to mid Hauterivian.
Environment: Neritic, carbonate substrate; higher numbers of attached forms such as Patellina subcretacea Cushman & Alexander, Trocholina infragranulata Noth, Globospirillina neocomiana (Moullade), and Spirillina spp., indicate shallow neritic conditions.
Remarks: This assemblage displays high species diversity and high numbers of specimens. Preservation is good, colour is reddish.
Occurrence: Found in the 33/9-15 well between 2731.7 and 2733 m, and observed in numerous other wells in the Viking Graben. The foraminiferal assemblage at the base of the studied section in well 33/9-15 is dominated by Globospirillina neocomiana and Patellina subcretacea. At 2731.7 m, Falsogaudryinella praemoesiana is still common, but in the lower sample (2733 m) it is only represented by a single specimen. Accessory species include Lenticulina munsteri, ammodiscids, and nodosariids. A single occurrence of a species of Protomarssonella that resembles M. kummi is present at 2733 m.
This type of assemblage has not been observed offshore mid-Norway, where the lower part of the Lower Cretaceous interval (between the Upper Jurassic Spekk Formation and the Lower Cretaceous Lyr Formation) is missing.
Correlation: The assemblage probably corresponds to the Marssonella kummi (= FCN 3) zone of King et al. (1989) in the Viking Graben. The species M. kummi has been subsequently transferred to the genus Protomarssonella Banner & Desai (1987). This zone can be difficult to recognise, because the nominate species may be rare or absent.
The Trocholina infragranulata (=FCN 2) zone of King et al. (op.cit) was not utilized in this study, because its occurrence is facies-related, and clearly diachronous. King et al. regarded it as Valanginian, corresponding to the "basal Valhall limestone" facies of the Viking Graben. However, the species also occurs in limestones as young as Barremian in condensed facies deposited in shallower parts of the Viking Graben. Offshore mid Norway specimens were observed in the lower Barremian. Neagu (1975) also reported it from the Barremian of Romania.
None of the (more tethyan) shelly microfossil taxa reported for Hauterivian listed in the Millennium Atlas (Figure 2.4) were observed in Zone NCF1.
Falsogaudryinella praemoesiana Zone = NCF2 Zone
This zone contains a distinctive assemblage, defined on the concurrence of abundant Falsogaudryinella praemoesiana, abundant nodosariids and ostracods, and rare to common Patellina and Globospirillina (Figure 2.7a). In the study area, the zone corresponds to the basal part of the Lyr Formation. The following taxa were observed:
Falsogaudryinella praemoesiana LCO (abundant)
Falsogaudryinella tealbyensis (rare)
Falsogaudryinella xenogena (rare)
Recurvoides spp.
Glomospira spp. (rare)
Trocholina infracretacea (rare)
Patellina subcretacea
Globospirillina neocomiana
common to abundant nodosariids, and ostracods LCO
Lingulina semiornata LO
Lenticulina aff. subalata
Lenticulina cf. busnardoi LO
Lenticulina munsteri (common)
Lenticulina heiermanni LO
Lanticulina ouachensis wisselmanni LO(rare)
Citharina harpa (rare)
Planularia crepidularis (rare)
Epistomina tenuicostata LO (rare)
Epistomina hechti
Epistomina caracolla
Conorboides lamplughi LO
Lagena hauteriviana LO
Gavelinella sigmoicostata LO (rare)
small planktonics, including Blefusciana spp. (rare)
Age: Generally late Hauterivian, though locally persisting in early Barremian
Environment: Shallow to deep neritic.
Remarks: The upper boundary of the zone is recognised based on the LCO of Falsogaudryinella praemoesiana.The nominate species is abundant within the zone (as much as 50% of the assemblage), but it also persists in low numbers into the overlying zone. The assemblage is characterised by high species diversity, but also high dominance. It is often dominated by the species Falsogaudryinella praemoesiana, but Recurvoides and/or Glomospira may also be present in large numbers. The proportion of agglutinated taxa is higher compared with the underlying zone.
Direct comparisons with the type specimens of Falsogaudryinella moesiana Neagu described from the middle Albian of Romania revealed that the nominate species of this acme zone in the North Sea is indeed a different species. Because it has characters that are considered to be more primitive, Kaminski, Neagu and Platon (1995) erected the new species Falsogaudryinella praemoesiana.
Occurrence: The NCF2 Zone is readily observed in wells, offshore mid Norway. In the 33/9-15 well a single core sample collected from 2729 m contains an assemblage that is completely dominated by Falsogaudryinella praemoesiana and Protomarssonella subtrochus. Additional taxa include common ammodiscids (Ammodiscus tenuissimus, Glomospira charoides, G. gordialis), Lenticulina munsteri, and Globospirillina neocomiana. Rare planktonic foraminifers are represented by Blefuscuiana spp.
Correlation: This assemblage corresponds to the Falsogaudryinella moesiana (= FCN 4) zone of King et al. (1989). King equated the top of this zone to the top of the Hauterivian. However, identification of the top of acme zones is subjective. In the Viking Graben, the maximum number of Falsogaudryinella praemoesiana, on average are found below the Munk Marl horizon, which is assigned to lower Barremian, but in some well sections, the acme of this species probably persists into Barremian. The species Falsogaudryinella praemoesiana also occurs in the upper Hauterivian of Romania (Dambovicioara Valley). В
Falsogaudryinella xenogena Zone = NCF3 Zone
This zone is defined on the upper part of the ranges of Falsogaudryinella xenogena, F. praemoesiana, Patellina, and Globospirillina (Figure 2.7a). Assemblages assigned to the zone typically contain the following taxa:
Falsogaudryinella praemoesiana LO (rare)
Falsogaudryinella xenogena LO (rare)
Recurvoides spp.
Glomospira spp. (rare)
Textularia bettenstaedti
Ammobaculoides carpathicus LO (rare)
Trocholina infracretacea LO (rare)
Patellina subcretacea LO
Globospirillina neocomiana LO
common to abundant nodosariids and ostracods
Gavelinella barremiana FO
Lenticulina munsteri (common)
Lenticulina aff. subalata
Lenticulina guttata
Lenticulina saxonica LO
Saracenaria spp.
several unilocular taxa
Epistomina hechti LO
Epistomina ornata LO
Epistomina caracolla LO
Conorotalites bartensteini LO (rare)
small planktonics (Blefuscuiana spp.) (rare)
Age: Early Barremian
Environment: Neritic
Remarks: The assemblage assigned to this zone is easily distinguished from the overlying non-calcareous (agglutinated) foraminiferal assemblages by the presence of diverse, red-stained lenticulinids and nodosariids. The nominate species, Falsogaudryinella xenogena is never common; it co-occurs with F. praemoesiana.
Occurrence: Found widespread offshore mid Norway, where it is characteristic of the reddish, calcareous sediments of the Lyr Formation.
Correlation: Corresponds to the “Falsogaudryinella sp. X (= FCN 5) zone of King et al. (1989). The species illustrated by King et al. (op. cit.) was named Falsogaudryinella xenogena by Kaminski, Neagu and Platon (1995), partly because the authors wished to preserve the letter "x" in the name. The name also refers to the fact that is it "foreign", (i.e. not observed in the Tethys region). In the Viking Graben, the nominate species mostly occurs in the lower to middle Barremian. It generally occurs below the Munk Marl horizon, though it is occasionally observed in or above it, partially overlapping in range with Gavelinella barremiana, the nominate species of the overlying zone.
In the absence of the nominate taxon, the LO of diverse, reddish lenticulinids can be used to recognize the top of the zone. Offshore mid Norway, the sediments immediately overlying this zone are non-calcareous claystones, and its benthic foraminiferal assemblage is stained green. At localities in the Viking Graben (UKCS Blocks 21 and 22) where the Barremian section is more complete, the first downhole occurrence of diverse lenticulinids, and a peak in their relative abundance correlates to a level just below the Munk Marl (ca. 20 - 40 m below).
Offshore mid - Norway, the lateral equivalent of the Munk Marl horizon was not observed in the logs, possibly owing to the presence of a hiatus. This horizon represents a regional dysaerobic facies in the North Sea, and contains a distinctive, white-stained benthic foraminiferal assemblage, with common Glomospira. The species Glomospira gaultina is well represented in the Munk Marl.
Several taxa of this and the previous zone are listed for Hautervian-Barremian strata in the Millennium Atlas (Figure 4).
Gavelinella barremiana Zone = NCF4 Zone
This zone is defined on the upper part of the range of Gavelinella barremiana (Figure 2.7a). Assemblages assigned to this zone typically contain the following taxa:
Gavelinella barremiana LO
Glomospira spp. (common)
Recurvoides spp. (common)
small specimens of Gyroidinoides aff. infracretaceus
Age: Barremian
Environment: Deep neritic
Remarks: The assemblage consists of small-size, reddish-stained specimens. An accessory zonal species, tentatively identified as Gyroidinoides aff. infracretaceus, differs from the typical Albian form in its much smaller size and in possessing fewer chambers.
Occurrence: The zone is rarely observed offshore mid-Norway, due to the common presence of a hiatus. A single core sample collected at 2725 m in well 33/9-15 in the northern Viking Graben, recovered a rich foraminiferal assemblage assigned to this zone. The sample consists almost entirely of red-coloured calcareous benthic foraminifers, dominated by the Barremian index species Gavelinella barremiana Bettenstaedt. This species is the index taxon of the upper Barremian G. barremiana zone (= FCN 6 of King et al., 1989). The sample also includes diverse nodosariids, and single specimens of Gyroidinoides and Ammodiscus.
Correlation: Gavelinella barremiana Bettenstaedt is a cosmopolitan species, known from the upper middle Barremian to upper Barremian of the northern North Sea, and from the upper Barremian to lower Aptian of northwest Germany, and offshore Canada. This zone is equivalent to the G. barremiana (= FCN 6) zone of King et al. (1989). In the Millennium Atlas the nominate taxon is listed from middle Barremian and Early Aptian.
Verneuilinoides chapmani Zone= NCF5 Zone
This zone is defined on the upper part of the ranges of Verneuilinoides chapmani, and Caudammina crassa (Figure 2.7a). Assemblages assigned to this zone consist mostly of green-stained, deeper water agglutinated foraminifers. However, in some wells, calcareous taxa may also be present with few specimens, especially at the base of the zone. The following agglutinated taxa have been observed:
Verneuilinoides chapmani LO
Caudammina crassa
Recurvoides spp. (common)
Glomospira sp. (common)
Cribrostomoides nonioninoides LCO (frequent)
Kalamopsis grzybowskii (rare)
Glomospira gaultina (rare)
Bathysiphon sp. (rare)
Rhabdammina, Rhizammina (few)
Age: Late Aptian – early Albian
Environment: Outer neritic – upper bathyal
Remarks: Taxonomic diversity is relatively high, and some deeper water taxa are present, including Kalamopsis grzybowskii, Bathysiphon sp., and Caudammina crassa. Glomospira gordialis is more common in the lower part of the zone. Increased number of tubular agglutinated forms such as Rhabdammina, Rhizammina, and Bathysiphon is interpreted as indicating deposition in deeper water. Dinoflagellate events present in this zone include the LO of Lithodinia stoveri.
Occurrence: Observed widespread, offshore mid Norway.
Correlation: The NCF5 Zone is equivalent to the Verneuilinoides chapmani (= FCN 8) zone of King et al. (1989). In the lower part of the zone, the ratio of Glomospira gordialis to Glomospira charoides increases considerably. This feature is often observed in the lower Albian of the Viking Graben, and probably denotes a more dysoxic environment. In the lowermost part of the zone, pyritized specimens of Lenticulina, and other calcareous benthics are occasionally observed. This is consistent with the observation by King et al. (1989), that calcareous benthic foraminifers sometimes "re-appear down section in the lowest part of the [V. chapmani] zone". King et al. (op.cit.) correlated the lowest part of zone FCN 8 with the uppermost Aptian. In the wells studied, the base of this zone lies disconformably upon the Barremian (mostly lower Barremian) Lyr Formation. Therefore, a regional hiatus that encompasses most of the Barremian and Aptian is deemed to be present. The hiatus corresponds to a change in lithology from reddish marls and limestones below, to greenish non-calcareous claystones above.
Recurvoides/Glomospira Zone= NCF6 Zone
This assemblage zone contains the following characteristic species:
Recurvoides spp. (acme)
Plectorecurvoides irregularis
Glomospira charoides (abundant)
Glomospira gordialis
Ammodiscus spp.
Uvigerinammina una (rare)
Cribrostomoides nonioninoides (rare)
Textularia spp. (rare)
Pseudobolivina variabilis
Gyroidinoides infracretaceus
Gavelinella intermedia
Age: Middle Albian
Environment: Upper bathyal
Remarks: The assemblage is strongly dominated by Recurvoides spp., and Glomospira spp. Other agglutinated taxa listed above often occur as single specimens. Calcareous benthic and planktonic forms are rare in this zonal interval, and may represent caving.
Occurrence: The zone is missing in many wells due to a widespread hiatus of Aptian through early middle Albian age; it occurs condensed in well 6406/2-2.
Correlation: This zone corresponds to the Recurvoides sp. (= FCN 10a) zone of King et al. (1989). В The species "Globigerinelloides" gyroidinaeformis has not been observed offshore mid-Norway. It is, however, known from the northern Viking Graben in well 33/9-15 at 2723m, dated latest early Albian. The core sample collected at 2723 m in that well contains an assemblage comprised mainly of calcareous benthic species and Hedbergella. The dominant (and most distinctive) taxon is "Globigerinelloides" gyroidinaeformis Moullade, a species that is probably a benthic form because of slight streptospiral coiling, a feature unusual for planktonics. Moreover, it is often found in assemblages where other well-known planktonic forms are absent. This distinctive species is often observed in acmes (floods), and defines the latest early Albian "Globigerinelloides" gyroidinaeformis (FCN 9) zone in the Viking Graben of King et al. (1989). The species is also known from the Grand Banks, offshore eastern Canada, and from the Vocontian Basin, France. This stratigraphic level has not been observed offshore mid Norway; either the species is ecologically excluded from the area or a late Early Albian hiatus is present in the offshore mid-Norway area.
Uvigerinammina una Zone - NCF7 Zone
This zone is defined on the upper part of the range of Uvigerinammina una, together with Ammoanita globorotaliaeformis (Plate 2.1). Assemblages assigned to this zone generally consist of diverse, green-stained deep-water agglutinated foraminifers, with few calcareous benthics present, and common to frequent planktonics, belonging to few species. The following taxa have been observed in the zone:
Uvigerinammina una LO (common)
Falsogaudryinella alta
Textularia bettenstaedti (rare)
Haplophragmoides spp.
Reophax troyeri LO
Ammoanita globorotaliaeformis LO
Cribrostomoides nonioninoides LO
Trochammina abrupta
Gyroidinoides infracretaceus
Gavelinella intermedia
Pleurostomella barrowsi LO
Glomospira charoides
Glomospira gordialis
Hedbergella planispira FCO
Hedbergella infracretacea (common)
Hedbergella delrioensis (rare)
Globigerinelloides bentonensis (rare)
Age: Late middle to early late Albian
Environment: Upper bathyal
Remarks: Assemblages assigned to this zone, typically are comprised of agglutinated, and few calcareous benthic taxa, and often common to frequent planktonics, mostly Hedbergella planispira. The nominate taxon, Uvigerinammina una, apparently is a boreal species restricted to the Norwegian Sea and North Sea region. It appears not to possess calcareous cement, hence it cannot be assigned to the genus Falsogaudryinella. Most likely it belongs in the genus Uvigerinammina, as originally reported by Burnhill & Ramsay (1981).
It has not been found in the Albian of the Kirchrode-1 Borehole (Jaroslaw Tyszka, personal communication), or in North Atlantic oceanic deposits. The acme of Uvigerinammina una itself often forms a narrow zone, but it is also found in low numbers in both the overlying and underlying zones. Additionally, the LO of Pseudonodosinella troyeri (= Reophax minuta of other authors) and the LO of Textularia bettenstaedti have been observed in this zone. Both events are reported to occur within the early Albian "R. minuta zone" (zone FCS 8), of King et al. in the southern North Sea.
According to our observations, Ammoanita (Trochammina) globorotaliaeformis has a tighter coil than the Paleocene A. ingerlisae Gradstein & Kaminski and, on average more (not fewer) chambers in the last whorl. In local well completion reports such specimens are referred to as Trochamina 'globorotaliaformis', and constitute a useful mid-upper Albian index species.
Dinoflagellate events present in his zone include Apteodinium grande LCO, and Lithosphaeridium arundum LO; Ovoidinium scabrosum and Endoceratium turneri range upwards through this zone.
The number of planktonic foraminifers locally increases stratigraphically upwards in Zone NCF7, reflecting a change from more restricted marine (dysaerobic) to more oxygenated, open marine conditions. A similar change was reported by King et al., (1989) from the northern North Sea, as reflected in a change from non-calcareous to calcareous claystone (top of Sola Formation).
Occurrence: Found in a majority of wells studied, offshore mid-Norway.
Correlation: This zone corresponds to the Falsogaudryinella sp 1 (FCN 10b) zone of King et al. (1989). In the Kirchrode-1 Borehole in the Lower Saxony Basin, the change from dominant Hedbergella planispira (below) to Hedbergella delrioensis (above) is observed within the early late Albian P. columnata calcareous nannofossil zone, and the Ticinella raynaudi planktonic foraminiferal zone of Randrianasolo and Anglada (1989; Weiss, 1997). The zone probably correlates to the inflatum ammonite zone.
Osangularia schloenbachi Zone = NCF8 Zone
This zone is defined on the upper part of the stratigraphic range of Osangularia schloenbachi. Sigmoilina antiqua, Falsogaudryinella alta, and Globigerinelloides bentonensis co-occur (Figure 2.7). Assemblages assigned to this zone typically contain the following taxa:
Osangularia schloenbachi LO (rare to common)
Gavelinella intermedia
Gyroidinoides infracretaceus
Sigmoilina antiqua
Lenticulina muensteri
Epistomina spinulifera LO
Valvulineria gracillima (rare)
Falsogaudryinella alta LO (rare)
Clavulina gaultina
Glomospira charoides (often common)
Ammodiscus tenuissimus
Hedbergella infracretacea LO (common)
Hedbergella delrioensis FCO
Hedbergella planispira (rare)
Globigerinelloides bentonensis FO
pink coloured Inoceramus prisms
Age: Late Albian
Environment: Upper bathyal
Remarks: This is a diverse assemblage, comprised of calcareous and agglutinated taxa, stained green. The species Osangularia schloenbachi occurs as an acme, and rare specimens are often found in the underlying zone. The species is known from the upper Albian of Romania, Poland, Germany and France, and probably originated in Tethys (Price, 1976). In addition to the nominate taxon, Gavelinella intermedia, Gyroidinoides infracretaceus, and lenticulinids are also common, as well as Glomospira charoides. Planktonic foraminifers are often abundant, and are strongly dominated by small specimens of Hedbergella delrioensis, probably the stratigraphic onset (FCO) of a regional bloom that ends in the H. delrioensis LCO zone, as discussed below.
Dinoflagellate taxa present in the zone include Lithosphaeridium conispinum LO, Lithodinium arundum LO, Ovoidinium scabrosum LCO and Apteodinium grande LO, marker taxa for upper Albian (Costa and Davey, 1989; E. Monteil, pers. comm., 1997).
In the RASC scaled optimum sequence of Figure 2.5 this zone is represented by a tight cluster of 5 events, and in the corresponding optimum sequence of Figure 2.5 by taxa at rank positions 85 - 90.
Occurrence: Found in several of the wells, offshore mid-Norway. Where O. schloenbachi is rare, recognition of the zone may be hampered by cavings from overlying, fossiliferous strata assigned to lower Cenomanian. An uppermost Albian hiatus in some of the wells, may truncate part of this zone, possibly related to the pronounced eustatic offlap cited by Hardenbol et al. (1993), and by Rohl & Ogg (1996) at the base of the inflatum zone. The hiatus is visualized by the large interfossil distance below A. grande, at the base of the RASC interval zonein Figure 2.5.
Correlation: This zone corresponds to the Osangularia schloenbachi (= FCN 12a) zone of King et al. (1989; see also Crittenden, 1983). King et al. (1989) reported that the stratigraphical range of this species is diachronous from south to north, occurring in the mid-Albian in the southern sector of the North Sea, but persisting to the upper Albian in the Viking Graben. The underlying Globigerinelloides bentonensis (=FCN 11) zone of King et al. (1989) is not recognised in the study area owing to the rarity of this species. In the Kirchrode-1 Borehole in the Lower Saxony Basin, NW Germany, the level with abundant Globigerinelloides bentonensis correlates to the late Albian E. turriseiffelii calcareous nannofossil zone (Weiss, 1997). The zone probably correlates to the dispar ammonite zone.
Sigmoilina antiqua Zone = NCF9 ZoneВ В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В В
This zone is defined on the upper part of the stratigraphic range of Sigmoiliona antiqua. Textularia sp. 1 Burnhill&Ramsay, Pseudotextularia cretosa, Recurvoides imperfectus and Plectorecurvoides alternans also occur in this zone. Assemblages assigned to this zone typically contain the following taxa:
Sigmoilina antiqua LO
Pseudotextulariella cretosa LO
Spiroplectinata annectens LO
Arenobulimina advena
Marssonella ozawai
Textularia sp. 1 Burnhill&Ramsay (1981) LO
Textularia chapmani
Textularia foeda
В Plectorecurvoides alternans LO
Recurvoides imperfectus LO
Hippocrepina depressa
Valvulineria gracillima
Hedbergella delrioensis FCO
Hedbergella planispira
Gavelinella sp. X (unpublished) LO
Gavelinella intermedia
Lingulogavelinella jarzevae
pink coloured Inoceramus prisms
Age: Early Cenomanian
Environment: Upper bathyal
Remarks: Sigmoilina antiqua, the nominate taxon is rare. Its stratigraphic range extends from the two underlying zones, Uvigerinammina una Zone and O. schloenbachi Zone, into this zone. Average LO events that co-occur in this zone are Pseudotextulariella cretosa, Marsonella ozawai, Arenobulimina advena, and other taxa listed above. Gavelinella sp. X , with its steep last chamber is probably a new form, and together with Textularia sp. 1 may be restricted to the northern North Sea and Norwegian Sea. When and where found with sufficient number of specimens, offshore Norway, both taxa might be formally described as new taxa. The planktonic foraminifers Hedbergella delrioensis and H. planispira locally are common in this zone; Globigerinelloides bentonensis occurs with few specimens, and is only common in the immediately underlying zone.
The dinoflagellates Ovoidinium verrucosum and Endoceratium turneri on average top in this zone, and in fact form the constituent zonal cluster in the scaled optimum sequence (Figure 2.5), which correlates to the optimum sequence (Figure 2.5) at rank positions 80 to 84.
According to King et al., (1989), the LO events of foraminifers in this zone are indicative of lower Cenomanian strata, in agreement with the upper part of the stratigraphic range of the dinoflagellate taxon Ovoidinium verrucosum (see Costa and Davey, 1992).
Occurrence: Widespread offshore mid-Norway. In sandy lithology (for example in gravity flows assigned to the BlГҐlange Formation), this zone may be difficult to detect. The lack of clustering of events assigned to this zone in the scaled optimum sequence (Figure 2.5) indicates considerable 'noise', including missing data, in the wells that penetrate this stratigraphic level. The zone may be identified in well 6507/2-3 near 3350m, and in well 35/3-4 near 3370m.
Correlation: This zone corresponds to the Sigmoilina antiqua (= FCN 12b) zone of King et al. (1989), and to the upper part of the deep marine assemblage zone with Plectorecurvoides alternans (Geroch & Nowak, 1984). The zone probably correlates to the lower part of the mantelli/cantianum ammonite zone.
Hedbergella delrioensis LCO Zone = NCF10 Zone
This distinctive zone is defined on the LCO of Hedbergella delrioensis; it may occur in floods, accompanied by the LCO of H. planispira (Figures 2.5 and 2.9). The following taxa have been
Hedbergella delrioensis LCO (common to abundant)
Hedbergella planispira LCO (common to frequent)
Hedbergella sigali (rare)
Whiteinella brittonensis FO (isolated occurrence, rare)В В
Praeglobotruncana praehelvetica FO (isolated occurrence, rare)
Gavelinella intermedia (rare)
Gavelinella cenomanica (rare)
Valvulineria gracillima (rare)
Lingulogavelinella jarzevae LO (rare)
Arenobulimina spp. (rare)
Glomospirella gaultina
Recurvoides imperfectus LO
Hippocrepina depressa LO
Plectorecurvoides spp. LO
Textularia sp. 1 LO (rare)
Textularia foeda (rare)
Age: Late early Cenomanian, possibly extending into early middle Cenomanian
Environment: Upper bathyal
Remarks: In many wells, including 35/3-5 between 3035 and 2930m, 6507/2-3 between 3300 and 3200m, and 6507/6-2 near 2950m, a distinctive assemblage is observed with Hedbergella delrioensis LCO (last common and last consistent occurrence, as observed uphole), in association with Gavelinella intermedia, G. cenomanica, Valvulinera gracillima, Lingulogavelinella jarzevae, Plectorecurvoides alternans, Recurvoides imperfectus, Hippocrepina depressa, Glomospirella gaultina, Ammophaeroidina sp 1 (RRI), Eggerellina mariei, Textularia foeda, and Tritaxia pyramidata. The majority of benthic taxa assigned to this zone are rare. Among planktonics Hedbergella planispira LCO occurs slightly down in this zone; H. sigali, Whiteinella brittonensis and Praeglobotruncana praehelvetica occur at few localities only in this zone, and are rare; in two wells (35/3-5 and 36/1-2) single specimens of Rotalipora cf. greenhornensis were reported.
Among dinoflagellates occur (average LO) Rhombodella paucispina, Fromea sp. 2, Epelidosphaeridia spinosa, Xiphophoridium alatum and Litosphaeridium siphoniphorum. The latter two taxa may extend in the overlying zone. The first (up well) occurrence of Heterosphaeridium difficile is in this zone (e.g. in well 6507/7-1 at 3506m), as reported by Bell & Selnes (1997). These authors regard this event to be close to the boundary of lower and middle Cenomanian, in good agreement with stratigraphic evidence cited below.
The dinoflagellate assemblage with E. spinosa, R. paucispina, X. alatum and L. siphoniphorum overlap ranges in lower Cenomanian strata (Costa and Davey, 1992). The benthic foraminifer L. jarvzevae in the southern North Sea ranges upwards into the lower Cenomanian (King et al., 1989). The possible presence in this zone of the planktonic foraminifer Rotalipora greenhornensis would indicate an early to middle Cenomanian age. Recurvoides imperfectus and Plectorecurvoides irregularis commonly occur in the older zones described above, and occasionally range upwards into this zone, together with Hippocrepina depressa. The upper range of these taxa Geroch and Nowak (1984) assigned to the Plectorecurvoides alternans Zone in the Carpathians extends into the lower part of Cenomanian. According to Prokoph (1997), the upper part of the LCO of H. planispira and H. delrioensis in NW Germany corresponds to the dixoni ammonite zone of late early Cenomanian age. We adhere to this correlation for offshore Norway
From the age of the underlying and immediately overlying zones, and the evidence cited above, we assign the Hedbergella delrioensis LCO zone to the upper part of the lower Cenomanian, possibly extending into middle Cenomanian (see below).
Occurrence: Widespread and distinctive, offshore mid-Norway.
Correlation: The zone overlaps with the lower part of the northern North Sea zone of H. brittonensis (= FCN 13) of King et al., (1989) containing abundant hedbergellids, assigned broadly to the middle part of Cenomanian. Offshore mid-Norway, the top of the zone is below or in sand of the basal Lysing Member. Since recognition of the zone is dependent on suitable, shaly lithology (the widespread gravity flow sands are not fossiliferous), the full stratigraphic extent of the zone remains to be determined. It probably incorporates a flooding horizon in the upper part of the lower Cenomanian, truncated by the mid-Cenomanian non-sequence (like the Rouen hardground of northern France), discussed below. В
Whiteinella brittonensis Zone = NCF11 Zone
This zone is defined on the lower part of the range of Whiteinella brittonensis, with rare Hedbergella spp. and Praeglobotrunacana spp. Also present may be Gavelinella intermedia, Valvulineria gracillima, and Glomospirella gaultina. Dicarinella and Marginotruncana taxa are not present in this zonal interval. The following taxa have been observed in assemblages assigned to this zone:
Hedbergella delrioensis (rare)
Hedbergella planispira (rare)
Clavihedbergella simplex LO (rare)
Whiteinella brittonensis FO
Gavelinella cenomanica LO (rare)
Gavelinella intermedia LO (rare)
Lingulogavelinella spp.
Valvulineria gracillima LO (rare)
Bulbobaculites problematicus
Glomospirella gaultina LCO
Uvigerinammina (pre-) jankoi
Ammosphaeroidina sp.
Eggerellina mariei LO
Age: Late middle to early late Cenomanian
Enviroment: Upper bathyal
Remarks: The interval from the level of the dinoflagelates Batioladinium jaegeri LO upwards to Heterosphaeridium difficile LCO, between optimum sequence positions 68 and 47, groups relatively tightly (Figure 2.5). This tight clustering in the scaled optimum sequence is the result of frequent cross-overs of the events in the wells.
A tight, lower subgroup in this interval, corresponding to rank positions 68 to 64 in the optimum sequence of Figure 2.5 is assigned to the W.brittonensis Zone, with the events listed above, some of which are scarce, or absent in the northern part of the region under study. The depth interval in wells that might be expected to contain this zone is often impoverished in fossils due to poor sample recovery from turbine drilling, widespread paleo watermass dysaerobia, stratigraphic hiatusess, or gravity flow sands. This zone and the immediately overlying one may be difficult to separate.
As far as may be determined, this zone containsrare Hedbergella delrioensis, H. planispira, Uvigerinammina (prae-) jankoi, Glomospirella gaultina, Gavelinella intermedia, G. cenomanica, Ammosphaeroidina sp., Eggerellina mariei, and Whiteinella brittonensis. Occasional events are Clavihedbergella simplex, and a specimen of Rotalipora cf. greenhornensis in southern well 36/1-2 at 2660m. The age assigned to the zone is late middle to early late Cenomanian.
Among dinoflagellate occur Batioladinium jaegeri LO, Endoceratium dettmaniae LO, E. ludbrookii LO, Dorocysta litotes, and Lithosphaeridium siphoniphorum LO (southern wells only). These events together are assigned a late middle to early late Cenomanian age (Costa and Davey, 1992), in agreement with the age assigned to the W. brittonensis Zone.
Occurrence: The zone is widespread, offshore Norway, but not easy to recognize, due to sandy facies.
Correlation: The W. brittonensis zone corresponds to the zone of the same name for the shale facies of the northern North Sea (King et al., 1989). It is possible that the upper part of the zone includes part of the unfossiliferous beds that occur as a result of widespread dysaerobia, or anoxia near the Cenomanian/Turonian boundary (see below).В
Dicarinella Zone = NCF12 Zone
This zone is defined on the presence of Dicarinella hagni-indica, D. imbricata, or D. primitiva. Other key taxa that may be present include Whiteinella inornata, W. paradubia, Praeglobotrunaca stephani LO, and Hedbergella hoelzi LO. The following taxa have been observed in assemblages assigned to this zone:
Dicarinella hagni - indica LO
Dicarinella imbricata LO
Dicarinella primitiva LO
Gaudryina filiformis (= Gerochammina lenis)В В
Hedbergella delrioensis LO (isolated specimens)
H.planispira LO (isolated specimens)
Hedbergella hoelzi LO
Praeglobotruncana stephani LO
Whiteinella brittonensis LO
Whiteinella inornata
Whiteinella paradubia
Marginotruncana marginata FO
Heterohelix globulosa (rare)
Bulbobaculites problematicus LO
Stensioeina humilis LO (rare)
Lingulogavelinella spp. (rare)
Gavelinella balthica (rare)
Rzehakina spp FO (incl. R. minima)
Caudammina ovuloides
Uvigerinammina jankoi (rare)
Psammosphaera spp. (frequent)
Allomorphina pyriformis LO
Allomorphina halli LO
C/T boundary gamma spike (more than one spike may be present)
pyritized small diatoms (frequent)
pyritized spumellarian radiolarians (rare)
Age: Latest Cenomanian - middle Turonian
Enviroment: Upper bathyal
Remarks: For the reasons, mentioned also with the immediately underlying zone, this zone maybe difficult to separate from the underlying one. Where siliciclastic sedimentation rates are high, with predominant gravity -flow sands, a typical Psammophaera/small pyritized diatom biofacies occurs, sprinkled with isolated H. delrioensis, H. planispira or Dicarinella. In few wells, particularly in the south where sand is less common, planktonic foraminifers are more diverse than in the immediately underlying or overlying zones, and include Praeglobotruncana stephani, P. delrioensis, Dicarinella hagni - indica, Dicarinella imbricata, Marginotruncana marginata FO, Dicarinella primitiva, Hedbergella hoelzi, H. delrioensis (rare), Whiteinella archeocretacea, W. inornata, W.paradubia, and W. brittonensis. Marginotruncana is mostly rare and only represented by M. marginata. From the overlap of planktonic foraminiferal ranges, the zone may be assigned a late Cenomanian through early middle Turonian age (Robaszynski et al., 1983; Stainforth, 1981). Marginotruncana marginata and Dicarinella are restricted to post Cenomanian strata and assist with constraining the age of the upper part of the zone.
It is likely that rare last occurrence in this zone of Uvigerinammina jankoi is in situ, since this taxon is known from the Turonian through Santonian (see e.g. Geroch and Nowak, 1984).
The assignment of an event labelled Cenomanian/Turonian (C/T) boundary gamma spike reflects our hypothesis that a distinct and narrow (less than a few meters thick) gamma ray high on physical well logs, near the base of this zone, corresponds to the well-known, global oceanic, 'anoxic', Event -2 (Bonarelli), reported widely in latest Cenomanian through earliest Turonian strata. The exact reason for the spike in the wells is unclear, but it is assumed to be high thorium concentration, associated with organic rich levels. We have no reason to believe that the spike is a local feature; in few wells more than one spike occurs over a short interval in this zone. From cuttings it cannot be ascertained that the spike is actually associated with an interval barren of microfossils, but in cores in well 6507/2-3 a distinct reduction to virtually no microfossils at and below the C/T boundary gamma spike at 2972m is observed. A peculiar, monotypic Nyktericysta / Australosphaera dinocyst assemblage maybe more or less associated with this 'Bonarelli' type interval (G. Bell, pers. comm., 1996), although RASC does not link the two events closely on average (Figure 2.5). Reason for the latter may be high variance in the average stratigraphic position of both events.
Considerable confusion exists over the identity of triserial to twisted biserial agglutinated benthic foraminifer specimens commonly found in the middle to upper Cretaceous of wells. Caved (?) Karrerulina conversa may be mislabelled as Gaudryina filiformis, and be difficult to distinguish from it. A detailed taxonomic study of such morphotypes in the wells is warranted.
In core sample 2885.5m in well 6507/2-3, assigned to this zone, Rzehakina minima has its first stratigraphic occurrence. Such would agree with the report by Kaiho and Hasegawa (1994) that in Japan Rzehakina (R. epigona) appears in Turonian strata. Bulbobaculites problematicus commonly occurs in this zone, and rarely in the immediately underlying one, which agrees with broad age assignment for this taxon in Kuhnt & Kaminski (1990).
Among dinocyst events occur Maghrebinia membraniphorum LO, Palaeohystrichosphera infusorioides acme and LCO, Heterosphaeridium difficile FCO, Stephodinium coronatum LO, and Spiniferites porosus. According to Costa and Davey (1992) these taxa are associated with Turonian strata. Dorocysta litotes LO alo may be present, but was omitted from RASC input data due to erratic stratigraphic occurrence.
The age of the Dicarinella zone is Turonian in its upper part, while the lower part may extend downward into Cenomanian, hence the zone is bracketed to occur across the C/T boundary. High sedimentation rates, a sandy, dysaerobic facies with occasionally much pyrite, and turbo drilling in many wells prevent satisfactory stratigraphic resolution across this interval.
Occurrence: This zone is widespread and relatively thick, offshore Norway.
Correlation: The Dicarinella zone corresponds to the zone of the same name in the northern North Sea, characterised by a downhole influx of abundant planktonic foraminifers, dominantly hedbergellids and Dicarinella spp. (King et al., 1989). The lower part of the zone contains strata coeval to the organic- rich Plenus marls and the BlГёdoks shales of the North Sea and England.
Marginotruncana Zone = NCF13
This zone is defined on the presence of Marginotruncana marginata, M. coronata, Globotruncana linneiana gr., and G. fornicata (primitive morphotype). The following taxa have been observed in assemblages assigned to this zone:
Margintruncana marginata
Marginotruncana fornicata (primitive morphotype)
Marginotruncana coronata
Globotruncana linneiana gr.
Dicarinella concavata (isolated specimens)
Dicarinella carinata (isolated specimens)
Whiteinella archeocretacea
Conorbina supracretacea
Lingulogavelinella sp.
Nuttalina florealis
Globorotalites spp.
Gaudryina filiformis
Uvigerina jankoi
Haplophragmoides aff. walteri (sensu Kuhnt & Kaminski, 1990)
Inoceramus needles
Age: Late Turonian - Coniacian
Enviroment: Upper bathyal
Remarks: This zone is recognized in few wells in the southern part of the study region, where it constitutes a downhole influx of Marginotruncana, Globotruncana and Whiteinella. A distinct influx of Marginotruncana, together with Whiteinella was for example observed in the 35/3-4 well, together with isolated specimens of Dicarinella concavata and D. carinata. The overlap of ranges of the latter two taxa is indicative of Coniacian strata (Robaszynski et al., 1983).
The zone constitutes the upper part of high sedimentation rate interval and sandy facies in northern wells, where globotruncaniid planktonic foraminifers are mostly absent; hence recognition of the zone may be difficult. The large interfossil zone limit in Figure 2.5 (left) at H. difficile LCO reflects this important stratigraphic facies change; in the optimum sequence of Figure 2.5 (right) the zone is recognized at rank positions 47 - 50.
The (local) relative abundance of Marginotruncana, together with H. difficile probably is the result of a condensed sedimentary sequence under open marine and relatively warm water conditition. Unfortunately, the dinoflagellate cyst Heterosphaeridium difficile may display more than one LCO or acme, hence this event may not be unique. The acme ccurrence of H. difficile, according to Costa and Davey (1992) suggests a Turonian age, with the taxon ranging into the Coniacian. Among dinoflagellate cyst that range through the upper part of the zone, and may extend slightly in the overlying foraminiferal S. granulata polonica zone, are Chatangiella sp. 1 Stratlab, Trithyrodinium 'reticulatum', Surculosphaeridium longifurcatum and Heterosphaeridium difficile
Occurrence: This zone is clearly recognized in southern well 35/3-4 from 2425 -2460m. Where globotruncaniid foraminifers are missing, as in many northern wells, recognition of the zone may be difficult.
Correlation: A M. marginata zone is defined by King et al., (1989) for the northern North Sea, and was assigned a late Turonian through early Coniacian age. These authors extend the overlying S. granulata polonica interval zone beyond the Santonian range of the nominate taxon down into the upper Coniacian. We restrict the S. granulata polonica zone to the Santonian.
Stensioeina polonica Zone = NCF14
This distinctive zone is defined on Stensioeina (granulata) polonica LO. The following taxa have been observed in assemblages assigned to this zone:
Stensioeina (granulata) polonica LO
Globorotalites multiseptus
Nuttalina florealis
Gavelinella beccariiformis FOВ В
Epistomina supracretacea
Gyroidinoides beisseli
Globotruncana marginata (rare)
Globotruncana linneiana group (rare)
Globotruncana fornicata (rare)
Heterohelix sp.
Inoceramus needles (common)
Archeoglobigerina spp.
Allomorphina sp.
Tritaxia spp.
Age: Early to middle Santonian
Enviroment: Upper bathyal
Remarks: The nominate zone marker is the key taxon in this easily recognizable zone, with the last stratigraphic occurrence of the nominate species delimiting its upper boundary. The zonal interval also contains various other calcareous taxa, listed above. Planktonic foraminifers decrease stratigraphically upwards in this zone, and are much more typical for the immediately underlying zone. Deep marine agglutinated benthics mostly are scarce, but less so offshore mid-Norway. Uvigerinammina jankoi only was observed in few southern wells. Lenticular radiolarians also occur. In some wells the immediately overlying Inoceramus LCO zone and this zone are merged (see below). В
Occurrence: This zone occurs widespread, offshore Norway, and in some wells where the Inoceramus LCO zone merges with this zone, constitutes the first downhole carbonate rich interval, below the thick siliciclast interval assigned a Campanian age.
Correlation: This zone correlates to the Stensioeina granulata polonica zone of Koch (1977) of early and early middle Santonian age in NW Germany. This is the only Stensioeina taxon from the evolutionary plexus of Stensioeina that is widespread, offshore Norway, and also is observed in the Voering Basin. Elsewhere in NW Europe six or more successive Stensioeina zones are recognized from Turonian to Campanian age. King et al. (1989) also recognizes the polonica zone (FCS18), but these authors extend the zone below the range of the nominate species down into the Coniacian.
Inoceramus LCO Zone = NCF15
This zone contains the following characteristic event:
Inoceramus LCO
This zone is recognized from a relative abundance of Inoceramus needles that constitute the first carbonate rich interval downhole following a thick, siliciclastic Campanian section, with pyritized diatoms, deep water agglutinated foraminifers, and locally radiolarians. In a few wells, such as 35/3-4 this zone merges with the immediately underlying S. granulata polonica zone, but on average the two events are clearly superpositional (Figure 2.5), possess low variances, and are assigned to successive and separate zones. Spherical and lenticular radiolarians may be common in this zone, but are more so in the overlying Fenestrella bellii zone. In the southern part of the study area (34 and 35 blocks), the lower stratigraphic ranges of Tritaxia dubia, Globotruncana bulloides and Whiteinella baltica are in this zone, and Archaeoglobigerina may also be present
The sudden disappearance of common Inoceramus needles, stratigraphically upwards suggests a regional hiatus to span a part of upper Santonian and lower Campanian.
Age: Late Santonian
Enviroment: bathyal
Occurrence: The Inoceramus LCO zone is widespread, offshore Norway, and easily recognizable.
Correlation: This zone correlates to the 'radiolaria' zone, or part of it, in the North Sea (King et al., 1989) which zone according to these authors may have a somewhat spotty distribution northward. The latter might be the result of local dissolution of silica. The distribution of the Inoceramus zone north of the North Sea reflects a temporal increase of carbonate productivity during a time of limited terrigenous sedimentation.
Fenestrella bellii Zone = NCF16
This distinctive zone is defined on the presence of a small, pillbox diatom with an oval cross-section, a sharp periphery and sometimes a small dimple, named Fenestrella bellii (Appendix 1 and Plate 1). It may occur in floods, accompanied by rare to common deep water agglutinated foraminifers, and rare to common spherical/lenticular radiolarians. The following taxa have been observed in assemblages assigned to this zone:
Fenestrella bellii LO
spherical/lenticular radiolarians (rare to common)
Rzehakina epigona LO
Haplophragmoides aff. walteri LO (sensu Kaminski&Kuhnt, 1990), (rare)
Spiroplectammina navarroana
Trochamminoides spp.
В В Deep water agglutinated foraminifers, undifferentiated
Age: Early Campanian
Enviroment: Dysaerobic, middle to upper bathyal
Remarks: Above the Inoceramus LCO zone, an interval occurs in many wells with a small, morphologically distinctive, pyritized diatom, informally named Diatom sp. H in well reports, and here assigned to Fenestrella bellii (see Appendix 1). Spherical/lenticular radiolarians may be common, and so are coarse agglutinated benthic foraminifers with organic cemented tests. The latter include Karrerulina conversa, Haplophragmoides aff. walteri (sensu Kaminski & Kuhnt, 1990), Spiroplectammina navarroana, Paratrochamminoides olszewska, P. mitratus, Trochamminoides subcoronatus, and Rzehakina epigona, all of which may extend slightly higher in the section, above this zone. Rzehakina epigona has a high variance, and is not a reliable correlation event; although offshore Norway it disappears on average in the F. bellii zone.
Various taxa of Trochamminoides, listed above, in the North Atlantic underwent an evolutionary radiation in the early Campanian, an 'event' clearly recorded offfshore Norway in flysch-type, bathyal strata. In exploration well reports, the distinct Trochamminoides taxa are commonly lumped under Paratrochamminoides irregularis. The appearance and local disappearance (or probably more correctly, the part of the stratigraphic range ofthegenus where its taxa become rare) is in Campanian strata.In deeper bathyal settings, like in the Voering Basin, Gaudryina filiformis occurs in this zone (cf. Geroch & Nowak, 1984), but maybe difficult to separate from the younger ranging Gerochammina lenis Neagu. In Figure 2.5 (left) the F. bellii assemblage forms a distinct interval zone, separated with a large interfossil distance from the immediately overlying zone of Tritaxia dubia that includes the dinoflagellate Trithyrodinium suspectum LCO at its base.
Other dinoflagellate cyst taxa that have their average last occurrence in this zone include Paleoglenodinium cretaceum, while Chatangiella niiga, C. granulifera, Xenascus ceratoides and Trithyrodinium suspectum LCO on average disappear in the lowermost part of the immediately overlying zone (Figure 2.5).
Occurrence: Widespread offshore Norway, including the Voering Basin.
Correlation: This zone probably largely correlates to the 'Unnamed' zone FCN18 of King et al. (1989) with exclusively non-calcareous agglutinated foraminifers in the northern North Sea, assigned to lower Campanian, in agreement with our superpositional zone data and the upper range of the dinoflagellate cyst P. cretaceum in Costa and Davey (1992). In the southern North Sea, in UK and in NW Germany this stratigraphic interval is probably shallow marine, and hence dominated by carbonates. There it is zoned with Stensoeina taxa and planktonic foraminifers.
The lower Campanian Goesella rugosa zone of Geroch & Nowak (1984), first recognized in the Carpathian Trough, and based on the lower range of G. rugosa, is not recognized offshore Norway. The taxon is present in North Atlantic DSDP Sites and in Zumaia, Spain (Kuhnt and Kaminski, 1997).
The diatom, and locally radiolarian-rich interval in the Norwegian Sea wells, assigned to the F.bellii zone may correspond to the bio-silicious event in western Tethys and North Atlantic, observed in the upper part of magnetic anomaly 34 reversed (near the Santonian-Campanian boundary). This paleoceanographic event is a faunal change at a number of localities in the Atlantic and western Tethys (Kuhnt et al., 1989) coeval with a change from well-oxygenated reddish claystones below, to greenish, radiolarian-rich sediments above. This event has been termed Ocean Anoxic Event 3 (OAE 3), although there is no evidence for truly anoxic conditions at the seafloor. It reflects a time when surface productivity (and hence seafloor organic flux) was comparatively high.
Tritaxia dubia Zone = NCF17
This zone is defined on the presence of Tritaxia dubia LO, widely reported under a variety of names (see below). The following taxa have been observed in assemblages assigned to this zone:
Tritaxia dubia LO
Pseudogaudryina pyramidata LO (rare)
Marsonella crassa
Trochamminoides spp. LCO
Spiroplectammina dentata (rare)
Spiroplectammina navarroana (rare)
Spiroplectammina spectabilis
Arenoturrispirrilina sp. В
Caudammina ovulum
Caudammina ovuloides
Remesella varians
Glomospira spp.
Coarse agglutinated benthic foraminifers LCO
Epistomina supracretacea
Globorotalites michelinianus
Gyroidinoides beisseli LO
Gavelinella usakensis LO
Red coloured planktonic foraminifers
including Hedbergella, Rugoglobigerina, Marginotruncana, and Archeoglobigerina
Age: Middle to late Campanian
Enviroment: Upper bathyal
Remarks: In the southern part of the region studied, calcareous foraminifers may include abundant planktonics, as listed above, often stained red. Northward, calcareous foraminifers become rare, and the zone is more difficult to define, particularly when overlying Paleocene and Maastrichtian strata also contain coarse agglutinated foraminifers that may be caving. On average though, the latter disappear stratigraphically upward together with other taxa listed above, one of the most typical which is Tritaxia dubia. This taxon likely is a junior synonym of Tritaxia subparisiensis (Grzybowski) and in well reports may hide under junior synonyms like Pseudogaudryina capitosa, Tritaxia tricarinata and T. capitosa. Typically, several Spiroplectammina taxa may be present, including S. spectabilis, which regionally re-appears in the upper Paleocene, and Arenoturrispirillina.
The assemblage, together with the dinoflagellates listed below forms a distinctive interval zone in Figure 2.5 (left), which is assigned a middle to late Campanian age, based on the upper stratigraphic range of T. dubia (= T. subparisiensis) in Geroch and Nowak (1984).
Dinoflagellates are abundantly present, and include the average last occurrences of Trithyroidinium suspectum LCO, T. suspectum, T. castaneum, Chatangiella niiga, C. granulifera, C. ditissima, Spongodinium delitiense FO and Palaeohystrichophora infusorioides, which together belong in middle and upper Campanian (Costa and Davey, 1992). Using dinoflagellates, the LCO of T. suspectum and Chatangiella niiga LO may be used to distinguish a lower part of the T. dubia zone, middle Campanian.
Occurrence: Widespread and relatively thick, offshore Norway.
Correlation: After early Campanian, North Atlantic low diversity agglutinated foraminifer assemblages with Glomospira increase in diversity. New species belonging to the genera Caudammina, Haplophragmoides, Paratrochamminoides, and Rzehakina are observed for the first time, as are calcareous-cemented agglutinated forms belonging to the genera Arenobulimina, Clavulinoides, Dorothia, Goessella, Marsonella, Remesella, and Spiroplectinata. The latter assemblage change marks an Atlantic-wide drop in the level of the Carbonate Compensation depth (CCD) to a depth of below 5000 m, resulting in the deposition of the marly Crescent Peaks member of the Plantagenet Formation (Jansa et al., 1979). At this time, the distinctive ‘flysch-type’ and calcareous-cemented agglutinated taxa become common, and their overall diversity increase in the North Atlantic and also in the Norwegian Sea, reflected in the composition of the Tritaxia dubia zone. This faunal turnover coincides with major changes in watermass properties, owing to increased deep-water productivity from high-latitude sources, cooling and increased ventilation of deep waters, and higher upwelling rates. Active watermass communication through the Scotland-Greenland gateway (Figure 2.2) existed between the proto Norwegian Sea and the North Atlantic Ocean.
The Tritaxia dubia zone correlates to the Tritaxia capitosa zone, FCN19 of King et al (1989), also assigned to (middle to) upper Campanian. King et al (op cit.) also refers to the red claystones, assigned to the upper part of the zone, that stain the foraminifers red. The claystone corresponds to the 'Upper Red Unit' of the Flounder Formation in Moray Firth and 'Pink Chalk' in the central North Sea.
Reussella szajnochae Zone = NCF18
This zone is defined on the LCO of Reussella szajnochae. Other key events include Rugoglobigerina spp., Globigerinelloides volutus, and Globotruncanella havanensis. The following taxa have been observed in assemblages assigned to this zone:
Reussella szajnochae LO
Caudammina ovulum
Globotruncanella havanensis
Rugoglobigerina tradinghousensis (southern area only)
Rugoglobigerina rotundata
Rugoglobigerina spp.
Globotruncana arca (south only)
Globotruncana mariei (south only)
Globotruncanella intermedia
Globigerinelloides volutus
Hedbergella spp., including H. holmdelensis
Bolivinoides draco miliaris (southern area only)
Deep water agglutinated foraminifers (more commonly found northward)
Brizalina ex gr. incrasssata
Anomalinoides velascoensis
Stensioeina pommerana
Gavelinella beccariiformis
Age: Probably mostly early Maastrichtian.
Enviroment: Deep neritic to upper bathyal
Remarks: The zone of Reussella szajnochae is easily recognizable from the average LO of its distinctive, nominate species, which generally is present with few specimens only in the wells samples examined, but occurs widespread. Associated calcareous benthic taxa and planktonics are listed above. In southern wells many more taxa occur, that require study. In a few of these southern wells, isolated specimens of the nominate taxon range in the overlying zone of Pseudotextularia elegans. It is not clear if R. szajnochae is in fact reworked or extends stratigraphically upward. A similar pattern was observed also by King et al. (1989), suggesting the taxon is in fact time transgressive southward.
In few wells, the upper part of the R.szajnochae zone shows a bloom of Globigerinelloides volutus which may constitute a stratigraphically useful subzone.
Dinocysts that have their average last occurrence in this zone include Odontochitina costata and Aquilapollenites spp (Figure 2.5), typical for lower Maastrichtian (Costa and Davey, 1992).
Occurrence: Widespread, offshore Norway.
Correlation: The R. szajnochae zone correlates to the lower part of zone of the same name (FCN20 p.p.) in the northern North Sea (King et al., 1989). In the Carpathian belt, sediments of the same age contain Caudammina ovulum gigantea, a middle bathyal to abyssal taxon not observed offshore Norway. The taxon is known from the North Atlantic (Kaminski & Kuhnt, 1990), and the Faeroer Basin (D. van den Akker, pers. comm., 1997).
Pseudotextularia elegans Zone = NCF19
This distinctive zone is defined on presence of Pseudotextularia elegans LO (Figure 2.5); other key taxa that may be present include Rosita contusa, Rugoglobigerina spp., Racemiguembelina varians, and Brizalina ex. gr. incrassata. The following taxa have been observed in assemblages assigned to this zone:
Pseudotextularia elegans
Abathomphalus mayaroensis
Rosita (Globotruncana) contusa
Rugoglobigerina rugosa (includes acme)
Globigerinelloides volutus (includes acme)
Pseudoguembelina excolata
Racemiguembelina varians
Stensioeina pommerana
Brizalina ex gr. incrassata
Heterohelix spp.
Hedbergella spp., include. H. monmouthensis
Remarks: As far as can be determined, this zone is largely missing in northern wells (together with Danian, lower Paleocene). Where it is present, it is rich in calcareous taxa, predominantly planktonics. The lower part of the zone harbours acmes or LCO levels of Rugoglobigerina rugosa and/or Globigerinelloides volutus which may constitute 50-90% of specimens observed. More study, particularly of cores is desirable to understand the successive records of events in this zone.
Abathomphalus mayaroensis, where observed does not seem to occur at the top of the zone, but rather at its base or even lower. It is not clear from the data if such occurrences are due to caving, or indeed represent an older record. Dinoflagellate cysts present in the zone include Cerodinium diebeli and Palynodinium grallator LO, indicative of upper Maastrichtian (Costa & Davey, 1992).
Age: Late Maastrichtian
Enviroment: Deep neritic to upper bathyal.
Occurrence: The P. elegans zone is largely recognized in the southern part of the area studied. We postulate that this hiatus is due to shoulder uplift during ‘break-up’, prior to the onset of Paleogene seafloor spreading in the Norwegian Sea.
Correlation: The P. elegans zone correlates to the North Sea wide P. elegans zone of King et al. (1989), and the Gavelinella danica and P. elegans zones of Koch (1977) in NW Germany, correlative to the junior and casimirovensis belemnite zones (Late Maastrichtian).
In the zonal description mention was made of last common occurrence events, acmes or blooms and possible ‘flooding horizons’ of selected non-keeled planktonic foraminifers like Hedbergella planispira, H.delrioensis, Globigerinelloides volutus and Rugoglobigerina rugosa.Such levels may have good correlation potential. What is insightful is to compare planktonic foraminifer ranges in the UK and NW Germany with offshore Norway (Figure 2.9).
The boreal acmes offshore Norway of mid Cretaceous taxa compare to similar acmes in NW Germany and UK (Prokoff, 1997; Hart, 1993). Also shown on Figure 2.9 are the standard ammonite and planktonic foraminiferal zonations, and the regional RASC zones.
The prominent Globigerinelloides bentonensis acme in NW Europe is barely recognizable offshore Norway, which area is likely peripheral to its distribution. On the other hand, the offshore Norway acmes of Hedbergella delrioensis and H.planispira correlate well, albeit with lower stratigraphic resolution due to lower resolution sampling in wells. Why hedbergellids dramatically decline abundance near the boundary of lower and middle Cenomanian is not well understood, but some lines of evidence bear on the issue.
Firstly, in the Norwegian 35 block area the abrupt reduction in hedbergellids, the H.delrioensis LCO event, is followed by a level with abundant, sideritic concretions. The latter suggest that a local hiatus or condensed interval is present above that level, probably of early Middle Cenomanian age. Although a regional sequence stratigraphic analysis is lacking, we postulate that the H.delrioensis LCO event is indicative of the top of a transgressive or highstand sequence interval, and that the subsequent hiatus or lag represents a relative sealevel rise after a sudden lowstand. Hence, we propose that this hiatus is coeval with the Rouen hardground complex in France, the mid-Cenomanian non-sequence in southern UK and the large hiatus at the boundary of lower and middle Cenomanian in NW Germany, e.g. in the Baddeckenstedt area (Hardenbol et al.,1993; Mutterlose et al., 1997).
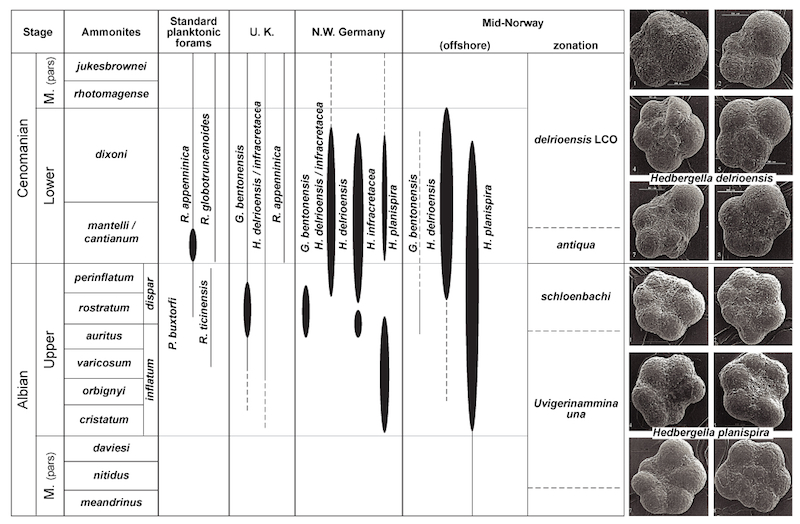
Figure 2.9. Boreal planktonic foraminifera acmes, offshore mid-Norway, correlated to similar acmes in NW Germany and U.K. (Prokoph, 1997; Hart 1993). Also shown are the standard ammonite and planktonic foraminifer zonations for the interval, and the regional RASC zonation. The prominent Globigerinelloides bentonensis acme in NW Europe is barely recognizable offshore Norway, which region is peripheral to its distribution. Reasons for the acmes are discussed in the text.
In this chapter we present a regional dinocyst zones (DCZ) with eleven zones and thirty-nine subzones for Cretaceous marine strata in the North Sea. HO stands for Highest (Stratigraphic) Occurrence and HCO for Highest Common (Stratigraphic) Occurrence, hence the zonation is optimized for industrial applications with ditch cuttings samples.
To increase utility of the operational micropaleontological zonation (discussed before; see also Figures 2.5 through 2.8) and the literature dinoflagellate zonations we have placed them side by side in Figures 2.10a-d, using their independent chronostratigraphic calibrations.
The chronostratigraphic interpretation of the dinoflagellate cyst events is mainly based on available peer-reviewed palynological publications. It also builds on results of unpublished PhD theses and collected contributions from the manuals of the “Advanced courses in Organic-Walled Dinoflagellate Cysts’’ organized from 1994 through 2012 in Utrecht, Haarlem, Bremen, Tubingen, Urbino and other locations. Use was made also of in-house (RGD, LPP and TNO) reports from 1980-2015 (e.g. TNO compilation report “Ranges and zonation of Cretaceous dinoflagellates in NW Europe, 2002” (see remarks below and reference list).
DCZ 1 Zone
Definition: The interval from the HCO of Cribroperidinium hansenii (preplicomphalus Ammonite Zone) to the HO of Daveya boresphaera (stenomphalus Ammonite Zone).
Age: Latest Volgian (latest Tithonian)-Berriasian
Remarks: Egmontodininium polyplacophorum has a HO in the oppressus Ammonite Zone (Partington et al., 1993) and the HO’s of Glossodiniium dimorphum (Riding and Thomas, 1992 and Partington et al., 1993) and Senoniasphaera jurassica (Riding and Thomas, 1992, Partington et al., 1993 and Poulsen, 1996) are in the anguiformis Ammonite Zone.
DCZ 1a subzone
Definition: The interval from the HCO of Cribroperidinium hansenii (preplicomphalus Ammonite Zone) to the HO of Egmontodinium expiratum (runctoni Ammonite Zone).
Age: Latest Volgian (latest Tithonian)-early Berriasian
Remarks: Davey (1982) reports the HO of Egmontodinium expiratum in the runctoni Ammonite Zone.
DCZ 1b subzone
Definition: The interval from HO of Egmontodinium expiratum (runctoni Ammonite Zone) to the HO of Systematophora daveyi (kochi Ammonite Zone).
Age:Middle Berriasian
Remarks: The HO of Systematophora daveyi is reported in the kochi Ammonite Zone by Davey (1992) and Poulsen (1996).
DCZ 1c subzone
Definition: The interval from HO of Systematophora daveyi (kochi Ammonite Zone) to the HO of Daveya boresphaera (stenomphalus Ammonite Zone).
Age: Late Berriasian
Remarks: The HO of Daveya boresphaera is defined by Davey (1979) and Birkelund et al. (1983). This taxon was better known as Gonyaulacysta sp. A and B sensu Davey (1979, 1982).
DCZ 2 Zone
Definition:The interval from the HO of Daveya boresphaera (stenomphalus Ammonite Zone) to the HO of Lagenorhytis delicatula (amblygonium Ammonite Zone).
Age:Valanginian
DCZ 2a subzone
Definition:The interval from the HO of Daveya boresphaera (stenomphalus Ammonite Zone) to the HO of Dingodinium spinosum (albidum Ammonite Zone).
Age:Earliest Valanginian
Remarks: The HO of Dingodinium spinosum in the albidum Ammonite Zone is published by the work of Duxbury (1977), Costa and Davey (1992) and Partington et al. (1993).
DCZ 2b subzone
Definition:The interval from the HO of Dingodinium spinosum (albidum Ammonite Zone) to the HO of Systematophora palmula (Polyptychites Ammonite Zone).
Age: Early Valanginian
Remarks: Reference for the HO of Systematophora palmula in the Polyptychites Ammonite Zone is Davey (1982) and Costa and Davey (1992).
DCZ 2c subzone
Definition:The interval from the HO of Systematophora palmula (Polyptychites Ammonite Zone) to the HO of Lagenorhytis delicatula (amblygonium Ammonite Zone).
Age:Late Valanginian
Remarks:The HO of Lagenorhytis delicatula in the amblygonium Ammonite Zone is reported in the compilation works of Heilmann-Clausen (1987) and Costa and Davey (1992).
DCZ 3 Zone
Definition: The interval from the HO of Lagenorhytis delicatula (amblygonium Ammonite Zone) to the HO of Cassiculosphaeridium magna (fissicostatum Ammonite Zone).
Age:Hauterivian-earliest Aptian
DCZ 3a subzone
Definition:The interval from the HO of Lagenorhytis delicatula (amblygonium Ammonite Zone) to the HO of Canningia cf. reticulata (lowermost variabilis Ammonite Zone).
Age:Early-middle Hauterivian
Remarks:The HO of Canningia cf. reticulata in lowermost variabilis Ammonite Zone is known from e.g. Duxbury (1977), Davey (1979), Costa & Davey (1992) and Heilmann-Clausen (1987).
DCZ 3b subzone
Definition:The interval from the HO of Canningia cf. reticulata (lowermost variabilis Ammonite Zone) to the HO of Nelchinopsis kostromiensis (variabilis Ammonite Zone).
Age: Late Hauterivian
Remarks: The HO of Nelchinopsis kostromiensis in the variabilis Ammonite Zone is referred to by Duxbury (1977; 2001) and Harding (1990).
DCZ 3c subzone
Definition: The interval from the HO of Nelchinopsis kostromiensis (variabilis Ammonite Zone) to the Kleithriasphaeridium corrugatum (elegans Ammonite Zone).
Age: Early Barremian
Remarks: The HO of Kleithriasphaeridium corrugatum in the elegans Ammonite Zone is reported in Duxbury (2001). Harding (1990) indicates the last occurrence in the fissicostatum Ammonite Zone of Speeton, UK, and in the Aulacoteuthis (belemnite) Zone of the Gott quarry in Germany. However, for the same quarry Heilmann-Clausen & Thomsen (1995) mention regular finds in the Upper Barremian brunsvicensis to lowermost germanica zones.
DCZ 3d subzone
Definition: The interval from the HO of Kleithriasphaeridium corrugatum (elegans Ammonite Zone) to the HO of Cassiculosphaeridium magna (fissicostatum Ammonite Zone).
Age: Late Barremian-earliest Aptian
Remarks: In the Speeton Clay this taxon is rare to fair up to bed LB2A, fissicostatum Zone. It is not recorded by Harding (1990) between LB1F (top fissicostatum Zone) and the basal part of the Upper B beds. Finds from this interval, which mainly includes the Middle B Beds, as mentioned by Davey (1974) and Duxbury (1980), are very rare. For the present study the HO of Cassiculosphaeridium magna is taken at the top of the fissicostatum Zone. Younger occurrences are rare and may (in part) be due to reworking; see also Mutterlose & Harding (1987), Heilmann-Clausen & Thomsen (1995), Lister & Batten (1995) and Below & Kirsch (1997).

Fig 2.10 partial: Comparison of the North Sea Dinocyst zonation with the Offshore Norway Foraminifer zonation. (Click for complete pdf)
DCZ 4 Zone
Definition: The interval from the HO of Cassiculosphaeridium magna (fissicostatum Ammonite Zone) to the HO of Dingodinium cerviculum (jacobi Ammonite Zone).
Age: Aptian
DCZ 4a subzone
Definition:The interval from the HO of Cassiculosphaeridium magna (fissicostatum Ammonite Zone) to the HO of Phoberocysta neocomica (forbesi Ammonite Zone).
Age:Earliest Aptian
Remarks:Duxbury (1983) shows a HO of Phoberocysta neocomica in the forbesi Ammonite Zone. In addition, the HO of Heslertonia heslertonensis is recorded in the forbesi Ammonite Zone by Duxbury (1983) and Lister & Batten (1988).
DCZ 4b subzone
Definition:The interval from the HO of Phoberocysta neocomica (forbesi Ammonite Zone) to the HO of Trichodinium ciliatum (bowerbanki Ammonite Zone).
Age:Early Aptian
Remarks:In England Lister & Batten recorded the taxon in the deshayesi and bowerbanki Zone. The HO of Callaiosphaeridium trycherium also verifies the top of the current subzone 4b (Duxbury, 1983).
DCZ 4c subzone
Definition: The interval from the HO of Trichodinium ciliatum (bowerbanki Ammonite Zone) to the HO of Muderongia sagena (nutfieldiensis Ammonite Zone).
Age: Middle Aptian
Remarks:The HO of Muderongia sagena in the nutfieldiensis Ammonite Zone is recorded by Lister & Batten (1988).
DCZ 4d subzone
Definition:The interval from the HO of Muderongia sagena (nutfieldiensis Ammonite Zone) to the HO of Dingodinium cerviculum (jacobi Ammonite Zone).
Age:Late Aptian
Remarks:The HO of Dingodinium cerviculum is uppermost jacobi Ammonite Zone (Duxbury, 1983). The top of the Dinocyst Zone 4 could also be ascertained by the HO of Gardodinium trabeculosum (Davey, 1982b).
DCZ 5 Zone
Definition:The interval from the HO of Dingodinium cerviculum (jacobi Ammonite Zone) to the HO of Cauca parva (dispar Ammonite Zone).
Age:Albian
DCZ 5a subzone
Definition:The interval from the HO of Cauca parva (dispar Ammonite Zone) to the HO of Discorsia nanna (tardefurcata Ammonite Zone).
Age:Earliest Albian
Remarks:According to Duxbury (1983) Discorsia nanna shows rare occurrences in the Upper Aptian and it becomes scarce in the tardefurcata Ammonite Zone. This last zone is also recorded by Costa & Davey (1992). In addition Occisucysta tentoria has its HO in the tardefurcata Ammonite Zone (Davey, 1982b and Duxbury, 1983).
DCZ 5b subzone
Definition:The interval from the HO of Discorsia nanna (tardefurcata Ammonite Zone) to the HO of Kleithriasphaeridium eoinodes (lautus Ammonite Zone).
Age:Early Albian
Remarks:The youngest occurrences of Kleithriasphaeridium eoinodes are mentioned by Verdier (1975) in the lautus Ammonite Zone. Davey (1982) shows the HO in the tardefurcata Ammonite Zone.
DCZ 5c subzone
Definition: The interval from the HO of Kleithriasphaeridium eoinodes (lautus Ammonite Zone) to the HO of Lithodinium arundum (inflatum Ammonite Zone).
Age:Middle-late Albian
Remarks: Lithodinium arundum has a HO in the inflatum Ammonite Zone (Davey & Verdier 1971, Fauconnier, 1975, Chateauneuf & Fauconnier, 1979, Foucher 1984) with a single occurrence in the topmost part of the dispar Zone (Tocher & Jarvis, 1996). Carpodinium granulatum could also be used for the top of the present subzone 5c (Verdier, 1975 and Foucher, 1983).
DCZ 5d subzone
Definition:The interval from the HO of Lithodinium arundum (inflatum Ammonite Zone) to the HO of Cauca parva (dispar Ammonite Zone).
Age:Late Albian
Remarks: Cauca parva is recorded in rare frequency in the uppermost Albian by Davey & Verdier 1973, Fauconnier, 1979 and Tocher & Jarvis, 1996. Younger Cenomanian occurrences (e.g. Foucher, 1984 and Prössl, 1990) are probably reworked (see discussion in Heilmann-Clausen, 1987).
The last consistent occurrence of Ovoidinium scabrosum is traditionally used for the top of the Albian (Gradstein, 2012). Ovoidinium scabrosum has an acme in the upper(most) Albian (TNO-NITG internal reports, Prössl, 1990). The LOD is lowermost Cenomanian (cantianum Zone, Foucher 1984, basal part varians Zone, Cookson & Hughes 1964, Davey 1970 and undifferentiated, Prössl 1990)
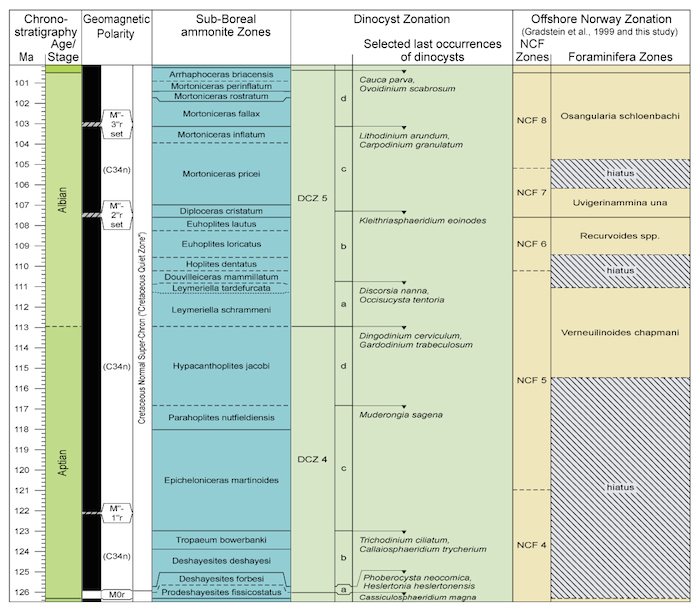
Fig 2.10 partial: Comparison of the North Sea Dinocyst zonation with the Offshore Norway Foraminifer zonation. (Click for complete pdf)
DCZ 6 Zone
Definition: The interval from the HO of Cauca parva (dispar Ammonite Zone) to the HO of Carpodinium obliquicostatum (judii Ammonite Zone).
Age: Cenomanian
DCZ 6a subzone
Definition:The interval from the HO of Cauca parva (dispar Ammonite Zone) to the HO of Protoellipsodinium spinocristatum (mantelli Ammonite Zone).
Age: Earliest Cenomanian
Remarks: Tocher & Jarvis (1996) mentioned the HO of Protoellipsodinium spinocristatum in the basal most part of the Lower Cenomanian, mantelli Ammonite Zone.
DCZ 6b subzone
Definition: The interval from the HO of Protoellipsodinium spinocristatum (mantelli Ammonite Zone) to the HO of Ovoidinium verrucosum (dixoni Ammonite Zone).
Age: Early Cenomanian
Remarks: Costa and Davey (1992) show a HO of Ovoidinium verrucosum in the dixoni Ammonite Zone. The top occurrence is confirmed by unpublished data of the Utrecht Knowledge Centre (LPP, TNO, RGD).
DCZ 6c subzone
Definition: The interval from the HO of Ovoidinium verrucosum (dixoni Ammonite Zone) to the HO of Epelidosphaera spinosa (guerangeri Ammonite Zone).
Age: Late Cenomanian
Remarks: The HO of Epelidosphaera spinosa is published by Tocher and Jarvis (1995) in the guerangeri Ammonite Zone. This age determination agrees with observations by Prössl (1990), who recorded the present taxon in regular to common values in the non-calibrated, basal Upper Cenomanian. Foucher (1979), Marschall (1983) and Costa & Davey (1992) show that last occurrence one ammonite zone older in the jukesbrownei Ammonite Zone.
DCZ 6d subzone
Definition:The interval from the HO of Epelidosphaera spinosa (guerangeri Ammonite Zone) to the HO of Carpodinium obliquicostatum (judii Ammonite Zone).
Age:Latest Cenomanian
Remarks: Regularly recorded up into the Plenus Marl (e.g. Clarke & Verdier, 1967, Davey, 1969, Foucher, 1979, 1980, 1983, Marshall 1983 and Dodsworth, 2000). Dodsworth (2000) reports the HO of Carpodinium obliquicostatum in the judii Ammonite Zone.
DCZ 7 Zone
Definition: The interval from the HO of Carpodinium obliquicostatum (judii Ammonite Zone) to the HO of Stephodinium coronatum (petrocoriensis Ammonite Zone).
Age: Turonian
DCZ 7a subzone
Definition:The interval from the HO of Carpodinium obliquicostatum (judii Ammonite Zone) to the HO of Codoniella campanulata (woollgari Ammonite Zone).
Age:Early Turonian
Remarks:The HO of Codoniella campanulata is reported by Pearce (PhD 2001) and may be associated with the woollgari Ammonite Zone.
В
DCZ 7b subzone
Definition:The interval from the HO of Codoniella campanulata (woollgari Ammonite Zone) to the HO of Stephodinium coronatum (petrocoriensis Ammonite Zone).
Age:Late Turonian
Remarks:The HO of Stephodinium coronatum in the petrocoriensis Ammonite Zone refers to Marschall (1983) and Schioler (1992).
DCZ 8 Zone
Definition:The interval from the HO of Stephodinium coronatum (petrocoriensis Ammonite Zone) to the HO of Scriniodinium campanula (socialis Zone)
Age:Coniacian-Santonian
DCZ 8a subzone
Definition:The interval from the HO of Stephodinium coronatum (petrocoriensis Ammonite Zone) to the HO (consistent) of Senoniasphaera rotunda alveolata (tridorsatum Ammonite Zone)
Age:Early Coniacian
Remarks:The interpretation of the HO (consistent) of Senoniasphaera rotunda alveolata in the tridorsatum Ammonite Zone is based on Pearce (2001) and Pearce et al. (2003).
DCZ 8b subzone
Definition: The interval from the HO of Senoniasphaera rotunda alveolata (tridorsatum Ammonite Zone) to the HO of Cyclonephelium filoreticulatum (serratomarginatus Ammonite Zone).
Age: Late Coniacian
Remarks: The interpretation of the HO (consistent) of Cyclonephelium filoreticulatum in the serratomarginatus Ammonite Zone is based on Prince et al. (1999) and Pearce (2001).
DCZ 8c subzone
Definition:The interval from the HO of Cyclonephelium filoreticulatum (serratomarginatus Ammonite Zone) to the HO of Psaligonyaulacysta deflandrei (polyopsis Ammonite Zone).
Age:Early Santonian
Remarks:The HO (consistent) of Psaligonyaulacysta deflandrei is in the Coniacian (Schioler 1992). In rare occurrences the taxon shows a HO in the early Santonian, polyopsis Ammonite Zone (Pearce, 2001).
DCZ 8d subzone
Definition:The interval from the HO of Psaligonyaulacysta deflandrei (polyopsis Ammonite Zone) to the HO of of Scriniodinium campanula (socialis Zone).
Age:Late Santonian
Remarks:The HO of Scriniodinium campanula is late Santonian, Uintacrinus socialis Zoneaccording Prince et al. (1999).

Fig 2.10 partial: Comparison of the North Sea Dinocyst zonation with the Offshore Norway Foraminifer zonation. (Click for complete pdf)
DCZ 9 Zone
Definition:The interval from the HO of Scriniodinium campanula (socialis Zone) to the HO of Callaiosphaeridium asymmetricum (phaleratum Ammonite Zone).
Age:Early Campanian
DCZ 9a subzone
Definition: The interval from the HO of Scriniodinium campanula (socialis Zone) to the HO (consistent) of Coronifera oceanica (basal bidorsatum Ammonite Zone).
Age:Earliest Campanian
Remarks:Regular and common occurrence of Coronifera oceanica is recorded up to the basal part of the Lower Campanian (O. pilula, Prince et al., 1999; and Pearce, 2001).
DCZ 9b subzone
Definition:The interval from the HO (consistent) of Coronifera oceanica (basal bidorsatum Ammonite Zone) to the HO of Chatangiella manumii (bidorsatum Ammonite Zone).
Age:Early Campanian
Remarks:The HO of Chatangiella manumii is shown in the bidorsatum Ammonite Zone by Pearce, 2001 and in unpublished compilations hereafter (e.g. “Advanced course in organic-walled dinoflagellate cysts, 2012; HO at 82.2 Ma” ).
DCZ 9c subzone
Definition: The interval from the HO of Chatangiella manumii (bidorsatum Ammonite Zone) to the HO (consistent) of Dinogymnium denticulatum (“mid” phaleratum Ammonite Zone).
Age: Early Campanian
Remarks: Pearce (2001,and in more recent unpublished compilations, see above) shows a HO (consistent) of Dinogymnium denticulatum in the “mid” phaleratum Ammonite Zone. Slightly older HO in the Santonian is given by Prince et al., 1999 and Clarke & Verdier, 1967). Irregular younger occurrences in the Maastrichtian, are mentioned by Wilson, 1974; Herngreen et al., 1986 and Kirsch, 1991).
DCZ 9d subzone
Definition: The interval from the HO (consistent) of Dinogymnium denticulatum (“mid” phaleratum Ammonite Zone) to the HO of Callaiosphaeridium asymmetricum (top of phaleratum Ammonite Zone).
Age:Early-mid Campanian
Remarks: The HO of Callaiosphaeridium asymmetricum at the top of phaleratum Ammonite Zone is interpreted from records in the Lower Campanian Vaals Formation and in the succeeding Zeven Wegen Member (Foucher 1983b) and fairly regular records up to the Lower- basal Middle Campanian elevata Zone and an isolated occurrence in the Middle Campanian ventricosa Zone (Kirsch, 1991). These data confirm the top at the boundary “Lower/Upper” Campanian in the compilation of Costa & Davey (1992) and Van Adrichem Boogaert & Kouwe, 1993-1997.
DCZ 10 Zone
Definition: The interval from the HO Callaiosphaeridium asymmetricum (phaleratum Ammonite Zone) to the HO of Trichodinium castanea (basal P. neubergicus/A.tridens Ammonite Zone).
Age: Late Campanian-earliest Maastrichtian
DCZ 10a subzone
Definition: The interval from the HO of Callaiosphaeridium asymmetricum (phaleratum Ammonite Zone) to the HO of Raetiaedinium truncigerum (polyplocum Ammonite Zone).
Age: Late Campanian
Remarks: The HO of Raetiaedinium truncigerum is recorded in the polyplocum Ammonite Zone by Pearce, 2001 and in unpublished compilations hereafter (e.g. “Advanced course in organic-walled dinoflagellate cysts, 2012).
DCZ 10b subzone
Definition: The interval from the HO of Raetiaedinium truncigerum (polyplocum Ammonite Zone) to the HO of Senoniasphaera rotunda (hyatti Ammonite Zone).
Age:Late Campanian
Remarks: Senoniasphaera rotunda is (very) common in the upper Turonian (FitzPatrick, 1995), and Santonian to Lower Campanian (Clarke & Verdier, 1967; Prince et al., 1999) and has a consistent HO in the late Campanian (Pearce, 2001 and later unpublished compilations, see above). Scarce occurrences are reported from the Maastrichtian (Wilson, 1974, Foucher, 1983 and Schioler et al., 1997).
DCZ 10c subzone
Definition: The interval from the HO of Senoniasphaera rotunda (hyatti Ammonite Zone) to the HO of Trichodinium castanea (basal P. neubergicus/A.tridens Ammonite Zone).
Age: Latest Campanian-earliest Maastrichtian
Remarks: The HO of Trichodinium castanea in the transitional latest Campanian/earliest Maastrichtian is based on Foucher, 1983b, 1985; Kirsch, 1991 and Herngreen, 1996).
DCZ 11 Zone
Definition: The interval from the HO of Trichodinium castanea (basal P. neubergicus/A.tridens Ammonite Zone) to the HO of Dinogymnium spp. (un-named interval).
Age: Maastrichtian
DCZ 11a subzone
Definition: The interval from the HO of Trichodinium castanea (basal P. neubergicus/A.tridens Ammonite Zone) to the HO of Odontochitina costata (P. neubergicus/A.tridens Ammonite Zone).
Age: Early Maastrichtian
Remarks: The HO of Odontochitina costata in the P. neubergicus/A.tridens Ammonite Zone is based on Wilson, 1974, Foucher, 1983b, 1985, Herngreen, 1998 and Van Adrichem Boogaert & Kouwe, 1993-1997.
DCZ 11b subzone
Definition: The interval from the HO of Odontochitina costata (P. neubergicus/A.tridens AmmoniteZone) to the HO of Apteodinium deflandrei (basal gollevillensis AmmoniteZone).
Age: Early Maastrichtian
Remarks: The HO of Apteodinium deflandrei in the basal gollevillensis AmmoniteZone is shown by Van Adrichem Boogaert & Kouwe, 1993-1997 and by unpublished dinoflagellate cyst stratigraphy from the Utrecht dinocyst course manual, 2012). Several authors record a HO in the Campanian (Wilson, 1974, Foucher, 1983a and 1983b, Neumann & Patel, 1983). Marheinecke (1992) and Brinkhuis & Schioler (1996) confirm occurrences in the Maastrichtian.
DCZ 11c subzone
Definition: The interval from the HO of Apteodinium deflandrei (basal gollevillensis AmmoniteZone) to the HO of Xenascus ceratoides (gollevillensis AmmoniteZone).
Age: Middle - Late Maastrichtian
Remarks: In their compilation Costa & Davey (1992) indicate occurrences up to the lowermost Upper Maastrichtian. Van Adrichem Boogaert & Kouwe (1993-1997) show the HO of Xenascus ceratoides in the late Maastrichtian (junior belemniteZone).
DCZ 11d subzone
Definition: The interval from the HO of Xenascus ceratoides (gollevillensis AmmoniteZone) to the HO of Dinogymnium spp.
Age: Late Maastrichtian
Remarks: The HO of Dinogymnium spp. on top of the Maastrichtian is deduced from Marheinecke (1992), Brinkhuis & Schioler (1996), Schioler et al. (1997), Herngreen et al. (1986), Wilson (1974), Kirsch (1991) and Van Adrichem Boogaert & Kouwe (1993-1997).
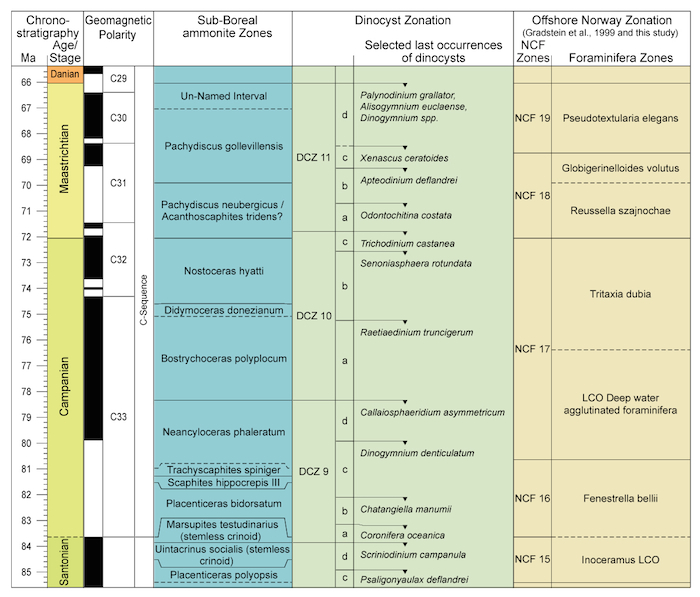
Fig 2.10 partial: Comparison of the North Sea Dinocyst zonation with the Offshore Norway Foraminifer zonation. (Click for complete pdf)